

Growth defect and mutator phenotypes of RecQ-deficient Neurospora crassa mutants separately result from homologous recombination and nonhomologous end joining during repair of DNA double-strand breaks
- PMID: 16219790
- PMCID: PMC1456140
- DOI: 10.1534/genetics.105.041756
Growth defect and mutator phenotypes of RecQ-deficient Neurospora crassa mutants separately result from homologous recombination and nonhomologous end joining during repair of DNA double-strand breaks
Abstract
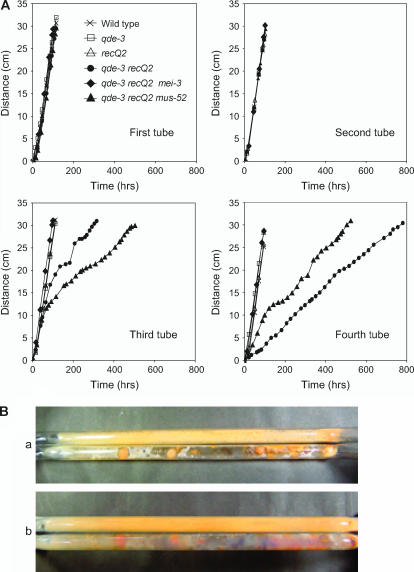
RecQ helicases function in the maintenance of genome stability in many organisms. The filamentous fungus Neurospora crassa has two RecQ homologs, QDE3 and RECQ2. We found that the qde-3 recQ2 double mutant showed a severe growth defect. The growth defect was alleviated by mutation in mei-3, the homolog of yeast RAD51, which is required for homologous recombination (HR), suggesting that HR is responsible for this phenotype. We also found that the qde-3 recQ2 double mutant showed a mutator phenotype, yielding mostly deletions. This phenotype was completely suppressed by mutation of mus-52, a homolog of the human KU80 gene that is required for nonhomologous end joining (NHEJ), but was unaffected by mutation of mei-3. The high spontaneous mutation frequency in the double mutant is thus likely to be due to NHEJ acting on an elevated frequency of double-strand breaks (DSBs) and we therefore suggest that QDE3 and RECQ2 maintain chromosome stability by suppressing the formation of spontaneous DSBs.
Figures






Similar articles
-
Srs2 and RecQ homologs cooperate in mei-3-mediated homologous recombination repair of Neurospora crassa.Nucleic Acids Res. 2005 Mar 30;33(6):1848-58. doi: 10.1093/nar/gki326. Print 2005. Nucleic Acids Res. 2005. PMID: 15800214 Free PMC article.
-
The Neurospora crassa mus-19 gene is identical to the qde-3 gene, which encodes a RecQ homologue and is involved in recombination repair and postreplication repair.Curr Genet. 2004 Feb;45(1):37-44. doi: 10.1007/s00294-003-0459-3. Epub 2003 Nov 1. Curr Genet. 2004. PMID: 14595518
-
Highly efficient gene replacements in Neurospora strains deficient for nonhomologous end-joining.Proc Natl Acad Sci U S A. 2004 Aug 17;101(33):12248-53. doi: 10.1073/pnas.0402780101. Epub 2004 Aug 6. Proc Natl Acad Sci U S A. 2004. PMID: 15299145 Free PMC article.
-
Mechanisms of DNA double strand break repair and chromosome aberration formation.Cytogenet Genome Res. 2004;104(1-4):14-20. doi: 10.1159/000077461. Cytogenet Genome Res. 2004. PMID: 15162010 Review.
-
The role of RecQ helicases in non-homologous end-joining.Crit Rev Biochem Mol Biol. 2014 Nov-Dec;49(6):463-72. doi: 10.3109/10409238.2014.942450. Epub 2014 Jul 22. Crit Rev Biochem Mol Biol. 2014. PMID: 25048400 Free PMC article. Review.
Cited by
-
Hitting the bull's eye: novel directed cancer therapy through helicase-targeted synthetic lethality.J Cell Biochem. 2009 Apr 1;106(5):758-63. doi: 10.1002/jcb.22048. J Cell Biochem. 2009. PMID: 19173305 Free PMC article. Review.
-
CaNRT2.1 Is Required for Nitrate but Not Nitrite Uptake in Chili Pepper Pathogen Colletotrichum acutatum.Front Microbiol. 2021 Jan 5;11:613674. doi: 10.3389/fmicb.2020.613674. eCollection 2020. Front Microbiol. 2021. PMID: 33469454 Free PMC article.
-
Neurosprora crassa RAD5 homologue, mus-41, inactivation results in higher sensitivity to mutagens but has little effect on PCNA-ubiquitylation in response to UV-irradiation.Curr Genet. 2007 Sep;52(3-4):125-35. doi: 10.1007/s00294-007-0146-x. Epub 2007 Aug 17. Curr Genet. 2007. PMID: 17703305
-
Human premature aging, DNA repair and RecQ helicases.Nucleic Acids Res. 2007;35(22):7527-44. doi: 10.1093/nar/gkm1008. Epub 2007 Nov 15. Nucleic Acids Res. 2007. PMID: 18006573 Free PMC article. Review.
-
Human RECQ1 is a DNA damage responsive protein required for genotoxic stress resistance and suppression of sister chromatid exchanges.PLoS One. 2007 Dec 12;2(12):e1297. doi: 10.1371/journal.pone.0001297. PLoS One. 2007. PMID: 18074021 Free PMC article.
References
-
- Bennett, R. J., J. A. Sharp and J. C. Wang, 1998. Purification and characterization of the Sgs1 DNA helicase activity of Saccharomyces cerevisiae. J. Biol. Chem. 273: 9644–9650. - PubMed
-
- Bennett, R. J., J. L. Keck and J. C. Wang, 1999. Binding specificity determines polarity of DNA unwinding by the Sgs1 protein of S. cerevisiae. J. Mol. Biol. 289: 235–248. - PubMed
-
- Brosh, R. M., Jr., D. K. Orren, J. O. Nehlin, P. H. Ravn, M. K. Kenny et al., 1999. Functional and physical interaction between WRN helicase and human replication protein A. J. Biol. Chem. 274: 18341–18350. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
