

High affinity ATP/ADP analogues as new tools for studying CFTR gating
- PMID: 16223764
- PMCID: PMC1464241
- DOI: 10.1113/jphysiol.2005.095083
High affinity ATP/ADP analogues as new tools for studying CFTR gating
Abstract
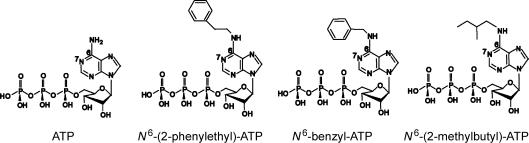
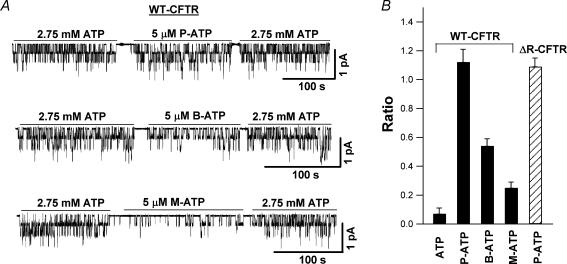
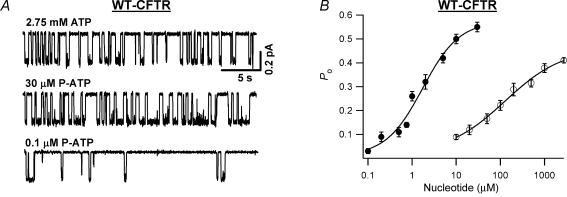
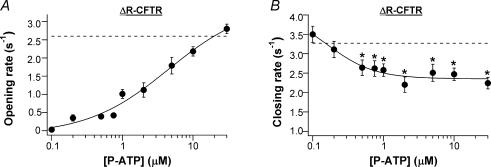
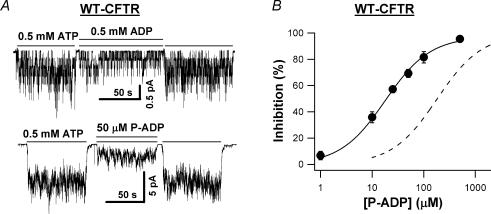
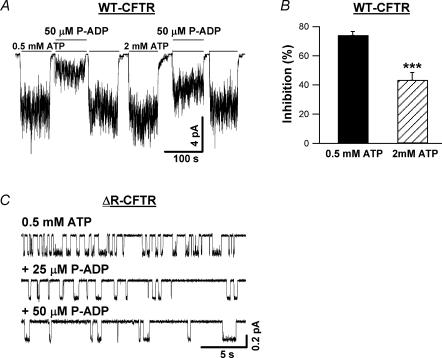
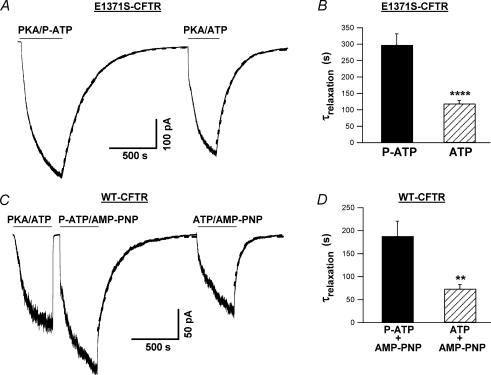
Previous studies using non-hydrolysable ATP analogues and hydrolysis-deficient cystic fibrosis transmembrane conductance regulator (CFTR) mutants have indicated that ATP hydrolysis precedes channel closing. Our recent data suggest that ATP binding is also important in modulating the closing rate. This latter hypothesis predicts that ATP analogues with higher binding affinities should stabilize the open state more than ATP. Here we explore the possibility of using N6-modified ATP/ADP analogues as high-affinity ligands for CFTR gating, since these analogues have been shown to be more potent than native ATP/ADP in other ATP-binding proteins. Among the three N6-modified ATP analogues tested, N6-(2-phenylethyl)-ATP (P-ATP) was the most potent, with a K(1/2) of 1.6 +/- 0.4 microm (>50-fold more potent than ATP). The maximal open probability (P(o)) in the presence of P-ATP was approximately 30% higher than that of ATP, indicating that P-ATP also has a higher efficacy than ATP. Single-channel kinetic analysis showed that as [P-ATP] was increased, the opening rate increased, whereas the closing rate decreased. The fact that these two kinetic parameters have different sensitivities to changes of [P-ATP] suggests an involvement of two different ATP-binding sites, a high-affinity site modulating channel closing and a low affinity site controlling channel opening. The effect of P-ATP on the stability of open states was more evident when ATP hydrolysis was abolished, either by mutating the nucleotide-binding domain 2 (NBD2) Walker B glutamate (i.e. E1371) or by using the non-hydrolysable ATP analogue AMP-PNP. Similar strategies to develop nucleotide analogues with a modified adenine ring could be valuable for future studies of CFTR gating.
Figures
References
-
- Ai T, Bompadre SG, Wang X, Hu S, Li M, Hwang TC. Capsaicin potentiates wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride-channel currents. Mol Pharmacol. 2004;65:1415–1426. - PubMed
-
- Aleksandrov AA, Aleksandrov L, Riordan JR. Nucleotide triphosphate pentose ring impact on CFTR gating and hydrolysis. FEBS Lett. 2002;518:183–188. - PubMed
-
- Aleksandrov L, Mengos A, Chang X, Aleksandrov A, Riordan JR. Differential interactions of nucleotides at the two nucleotide binding domains of the cystic fibrosis transmembrane conductance regulator. J Biol Chem. 2001;276:12918–12923. - PubMed
-
- Anderson MP, Welsh MJ. Regulation by ATP and ADP of CFTR chloride channels that contain mutant nucleotide binding domains. Science. 1992;257:1701–1704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous