

A paracrine peptide sex pheromone also acts as an autocrine signal to induce plasmid transfer and virulence factor expression in vivo
- PMID: 16223881
- PMCID: PMC1266105
- DOI: 10.1073/pnas.0505545102
A paracrine peptide sex pheromone also acts as an autocrine signal to induce plasmid transfer and virulence factor expression in vivo
Abstract
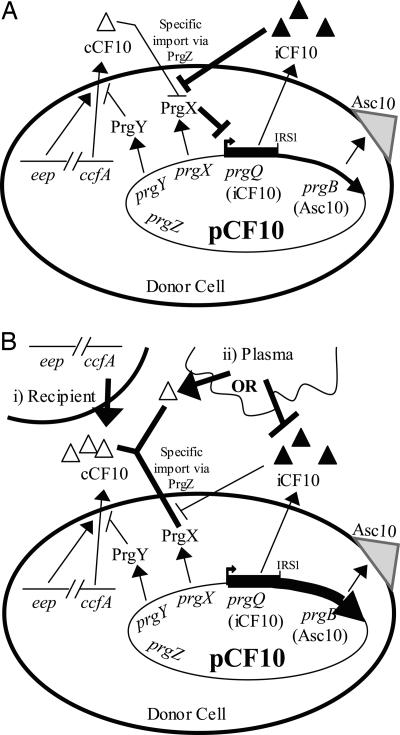
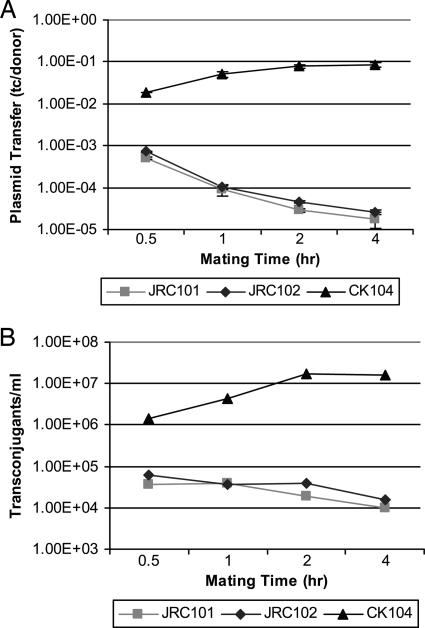
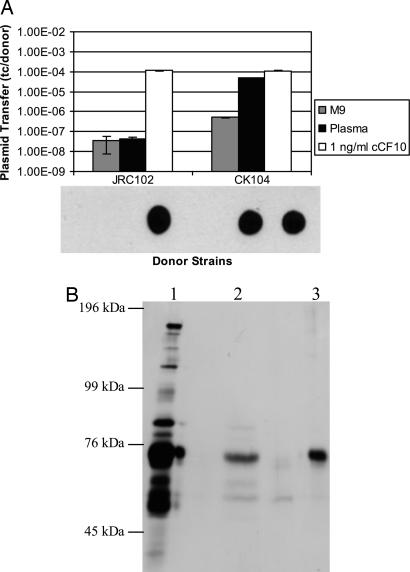
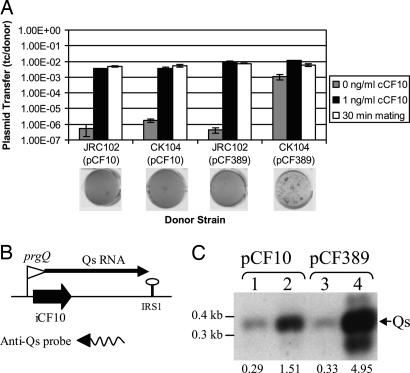
The peptide pheromone cCF10 of Enterococcus faecalis is an intercellular signal for induction of conjugative transfer of plasmid pCF10 from donor cells to recipient cells. When a donor cell is exposed to recipient-produced cCF10, expression of the pCF10-encoded aggregation substance of pCF10 (Asc10) and other conjugation gene products is activated. Asc10 also increases enterococcal virulence in several models, and when donor cells are grown in animals or in plasma, Asc10 expression is induced by means of the cCF10-sensing machinery. Plasmid pCF10 carries two genes that function to prevent self-induction by endogenous cCF10 in donor cells. The membrane protein PrgY reduces endogenous pheromone activity in donor cells, and the inhibitor peptide iCF10 neutralizes the residual endogenous cCF10 that escapes PrgY. In the current study, we found that E. faecalis strains with allelic replacements abolishing active cCF10 production showed reduced ability to acquire pCF10 by conjugation; prgY-null mutations had no phenotype in the cCF10-negative strains. We observed that expression of the mRNA for iCF10 was reduced in this background and that these mutations also blocked plasma induction of Asc10 expression. These findings support a model in which plasma induction in wild-type donors results from iCF10 inactivation by a plasma component, causing disruption of a precisely maintained balance of iCF10 to cCF10 activity and allowing subsequent induction by endogenous cCF10. Although cCF10 has traditionally been viewed as an intercellular signal, these results show that pCF10 has also adapted cCF10 as an autocrine signal that activates expression of virulence and conjugation functions.
Figures
Similar articles
-
Specific control of endogenous cCF10 pheromone by a conserved domain of the pCF10-encoded regulatory protein PrgY in Enterococcus faecalis.J Bacteriol. 2005 Jul;187(14):4830-43. doi: 10.1128/JB.187.14.4830-4843.2005. J Bacteriol. 2005. PMID: 15995198 Free PMC article.
-
Characterization of the sequence specificity determinants required for processing and control of sex pheromone by the intramembrane protease Eep and the plasmid-encoded protein PrgY.J Bacteriol. 2008 Feb;190(4):1172-83. doi: 10.1128/JB.01327-07. Epub 2007 Dec 14. J Bacteriol. 2008. PMID: 18083822 Free PMC article.
-
The prgQ gene of the Enterococcus faecalis tetracycline resistance plasmid pCF10 encodes a peptide inhibitor, iCF10.J Bacteriol. 1994 Dec;176(23):7405-8. doi: 10.1128/jb.176.23.7405-7408.1994. J Bacteriol. 1994. PMID: 7545961 Free PMC article.
-
The peptide pheromone-inducible conjugation system of Enterococcus faecalis plasmid pCF10: cell-cell signalling, gene transfer, complexity and evolution.Philos Trans R Soc Lond B Biol Sci. 2007 Jul 29;362(1483):1185-93. doi: 10.1098/rstb.2007.2043. Philos Trans R Soc Lond B Biol Sci. 2007. PMID: 17360276 Free PMC article. Review.
-
Peptide pheromone-induced transfer of plasmid pCF10 in Enterococcus faecalis: probing the genetic and molecular basis for specificity of the pheromone response.Peptides. 2001 Oct;22(10):1529-39. doi: 10.1016/s0196-9781(01)00489-2. Peptides. 2001. PMID: 11587782 Review.
Cited by
-
Enterococcus faecalis Sex Pheromone cCF10 Enhances Conjugative Plasmid Transfer In Vivo.mBio. 2018 Feb 13;9(1):e00037-18. doi: 10.1128/mBio.00037-18. mBio. 2018. PMID: 29440568 Free PMC article.
-
Pheromone-inducible conjugation in Enterococcus faecalis: a model for the evolution of biological complexity?Int J Med Microbiol. 2006 Apr;296(2-3):141-7. doi: 10.1016/j.ijmm.2006.01.040. Epub 2006 Feb 28. Int J Med Microbiol. 2006. PMID: 16503196 Free PMC article. Review.
-
Comprehensive Functional Analysis of the Enterococcus faecalis Core Genome Using an Ordered, Sequence-Defined Collection of Insertional Mutations in Strain OG1RF.mSystems. 2018 Sep 11;3(5):e00062-18. doi: 10.1128/mSystems.00062-18. eCollection 2018 Sep-Oct. mSystems. 2018. PMID: 30225373 Free PMC article.
-
Horizontal transfer of the tetracycline resistance gene tetM mediated by pCF10 among Enterococcus faecalis in the house fly (Musca domestica L.) alimentary canal.Microb Ecol. 2009 Oct;58(3):509-18. doi: 10.1007/s00248-009-9533-9. Epub 2009 May 28. Microb Ecol. 2009. PMID: 19475445
-
Enterococcal Sex Pheromones: Evolutionary Pathways to Complex, Two-Signal Systems.J Bacteriol. 2016 May 13;198(11):1556-1562. doi: 10.1128/JB.00128-16. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27021562 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
