

Unusual features of self-peptide/MHC binding by autoimmune T cell receptors
- PMID: 16226501
- PMCID: PMC3417822
- DOI: 10.1016/j.immuni.2005.09.009
Unusual features of self-peptide/MHC binding by autoimmune T cell receptors
Abstract
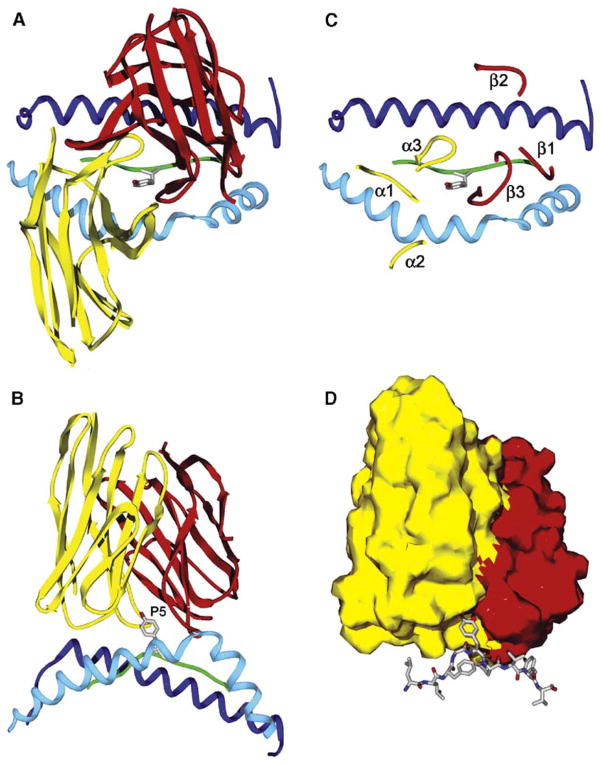
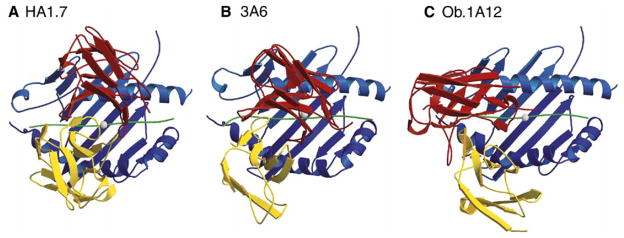
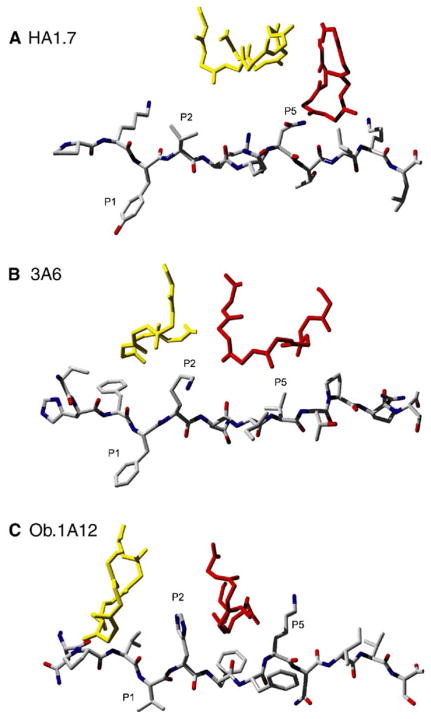
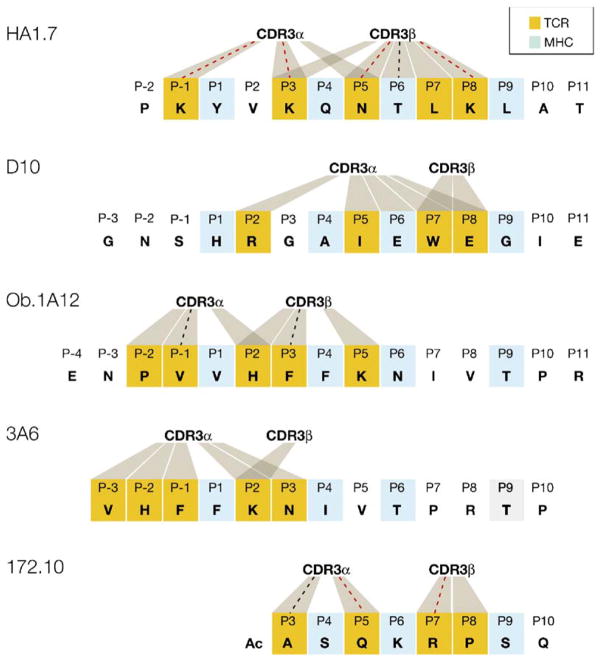
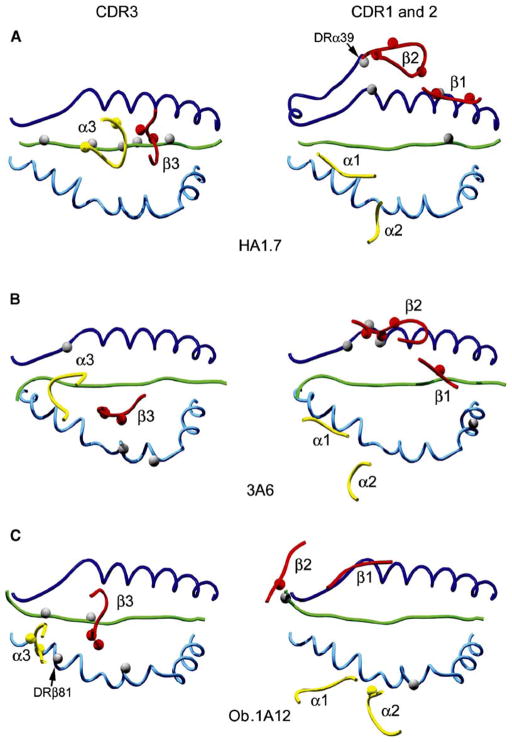
Structural studies on T cell receptors (TCRs) specific for foreign antigens demonstrated a remarkably similar topology characterized by a central, diagonal TCR binding mode that maximizes interactions with the MHC bound peptide. However, three recent structures involving autoimmune TCRs demonstrated unusual interactions with self-peptide/MHC complexes. Two TCRs from multiple sclerosis patients bind with unconventional topologies, and both TCRs are shifted toward the peptide N terminus and the MHC class II beta chain helix. A TCR from the experimental autoimmune encephalomyelitis (EAE) model binds in a conventional orientation, but the structure is unusual because the self-peptide only partially fills the binding site. For all three TCRs, interaction with the MHC bound self-peptide is suboptimal, and only two or three TCR loops contact the peptide. Optimal TCR binding modes confer a competitive advantage for antimicrobial T cells during an infection, whereas altered binding properties may permit survival of a subset of autoreactive T cells during thymic selection.
Figures
References
-
- Alam SM, Travers PJ, Wung JL, Nasholds W, Redpath S, Jameson SC, Gascoigne NR. T-cell-receptor affinity and thymocyte positive selection. Nature. 1996;381:616–620. - PubMed
-
- Anderson MS, Venanzi ES, Klein L, Chen Z, Berzins SP, Turley SJ, von Boehmer H, Bronson R, Dierich A, Benoist C, Mathis D. Projection of an immunological self shadow within the thymus by the aire protein. Science. 2002;298:1395–1401. - PubMed
-
- Appel H, Gauthier L, Pyrdol J, Wucherpfennig KW. Kinetics of T-cell receptor binding by bivalent HLA-DR. Peptide complexes that activate antigen-specific human T-cells. J Biol Chem. 2000;275:312–321. - PubMed
-
- Baker BM, Turner RV, Gagnon SJ, Wiley DC, Biddison WE. Identification of a crucial energetic footprint on the alpha1 helix of human histocompatibility leukocyte antigen (HLA)-A2 that provides functional interactions for recognition by tax peptide/HLA-A2-specific T cell receptors. J Exp Med. 2001;193:551–562. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
