

Reconfiguring the connectivity of a multiprotein complex: fusions of yeast TATA-binding protein with Brf1, and the function of transcription factor IIIB
- PMID: 16227432
- PMCID: PMC1266137
- DOI: 10.1073/pnas.0507653102
Reconfiguring the connectivity of a multiprotein complex: fusions of yeast TATA-binding protein with Brf1, and the function of transcription factor IIIB
Abstract
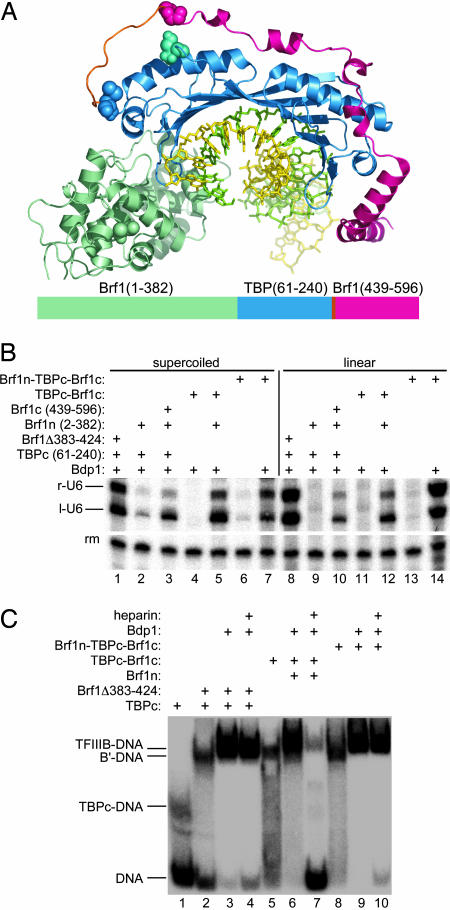
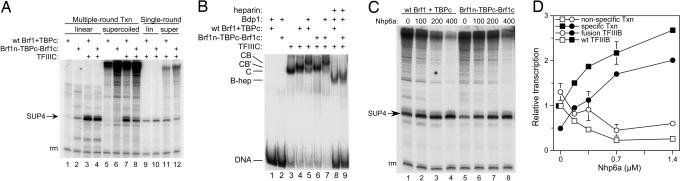
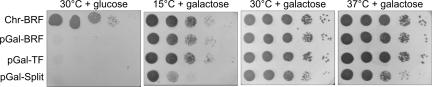
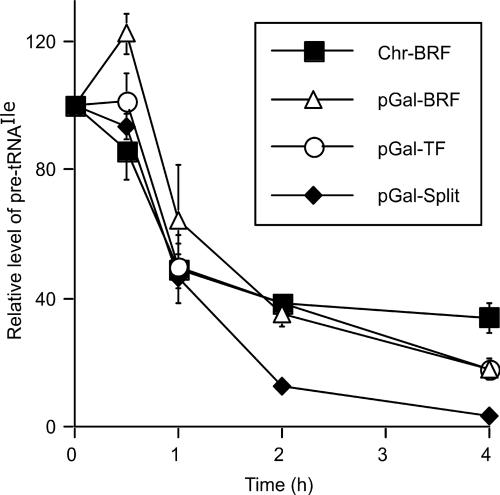
Transcription factor (TF) IIIB, the central transcription initiation factor of RNA polymerase III (pol III), is composed of three subunits, Bdp1, Brf1 and TATA-binding protein (TBP), all essential for normal function in vivo and in vitro. Brf1 is a modular protein: Its N-proximal half is related to TFIIB and binds similarly to the C-terminal stirrup of TBP; its C-proximal one-third provides most of the affinity for TBP by binding along the entire length of the convex surface and N-terminal lateral face of TBP. A structure-informed triple fusion protein, with TBP core placed between the N- and C-proximal domains of Brf1, has been constructed. The Brf1-TBP triple fusion protein effectively replaces both Brf1 and TBP in TFIIIC-dependent and -independent transcription in vitro, and forms extremely stable TFIIIB-DNA complexes that are indistinguishable from wild-type TFIIIB-DNA complexes by chemical nuclease footprinting. Unlike Brf1 and TBP, the triple fusion protein is able to recruit pol III for TATA box-directed transcription of linear and supercoiled DNA in the absence of Bdp1. The Brf1-TBP triple fusion protein also effectively replaces Brf1 function in vivo as the intact protein, creating a TBP paralogue in yeast that is privatized for pol III transcription.
Figures
References
-
- White, R. J. (2002) RNA Polymerase III Transcription (Landes Bioscience, Georgetown, TX).
-
- Hansen, S. K., Takada, S., Jacobson, R. H., Lis, J. T. & Tjian, R. (1997) Cell 91, 71-83. - PubMed
-
- Geiduschek, E. P. & Kassavetis, G. A. (2001) J. Mol. Biol. 310, 1-26. - PubMed
-
- Schramm, L. & Hernandez, N. (2002) Genes Dev. 16, 2593-2620. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
