

Functions and dysfunctions of the nuclear lamin Ig-fold domain in nuclear assembly, growth, and Emery-Dreifuss muscular dystrophy
- PMID: 16227433
- PMCID: PMC1255737
- DOI: 10.1073/pnas.0507612102
Functions and dysfunctions of the nuclear lamin Ig-fold domain in nuclear assembly, growth, and Emery-Dreifuss muscular dystrophy
Abstract
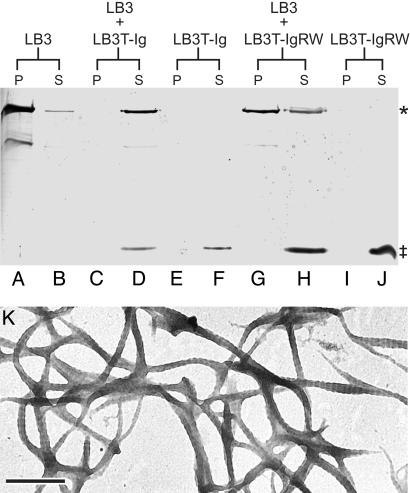
The non-alpha-helical C terminus of Xenopus lamin B3 (LB3T) inhibits the polymerization of lamin B3 in vitro and prevents the assembly of nuclei in Xenopus egg interphase extracts. To more precisely define the functions of LB3T in nuclear assembly, we have expressed subdomains of LB3T and determined their effects on nuclear assembly in Xenopus extracts. The results demonstrate that the Ig-fold motif (LB3T-Ig) is sufficient to inhibit lamin polymerization in vitro. Addition of the LB3T-Ig to egg extracts before the introduction of chromatin prevents chromatin decondensation and the assembly of the lamina, membranes, and pore complexes comprising the nuclear envelope. When added to assembled nuclei, LB3T-Ig prevents the further incorporation of lamin B3 into the endogenous lamina and blocks nuclear growth. The introduction of a point mutation in LB3T-Ig (R454W; LB3T-IgRW), known to cause Emery-Dreifuss muscular dystrophy when present in lamin A, does not inhibit lamin polymerization, chromatin decondensation, or nuclear assembly and growth. These results shed light on the specific alterations in lamin functions attributable to a known muscular dystrophy mutation and provide an experimental framework for revealing the effects of other mutations causing a wide range of laminopathies.
Figures




References
-
- Riemer, D., Wang, J., Zimek, A., Swalla, B. J. & Weber, K. (2000) Gene 255, 317-325. - PubMed
-
- Karabinos, A., Zimek, A. & Weber, K. (2004) Gene 326, 123-129. - PubMed
-
- Gruenbaum, Y., Margalit, A., Goldman, R. D., Shumaker, D. K. & Wilson, K. L. (2005) Nat. Rev. Mol. Cell Biol. 6, 21-31. - PubMed
-
- Hesse, M., Magin, T. M. & Weber, K. (2001) J. Cell Sci. 114, 2569-2575. - PubMed
-
- Strelkov, S. V., Schumacher, J., Burkhard, P., Aebi, U. & Herrmann, H. (2004) J. Mol. Biol. 343, 1067-1080. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
