

The splicing ATPase prp43p is a component of multiple preribosomal particles
- PMID: 16227579
- PMCID: PMC1265834
- DOI: 10.1128/MCB.25.21.9269-9282.2005
The splicing ATPase prp43p is a component of multiple preribosomal particles
Abstract
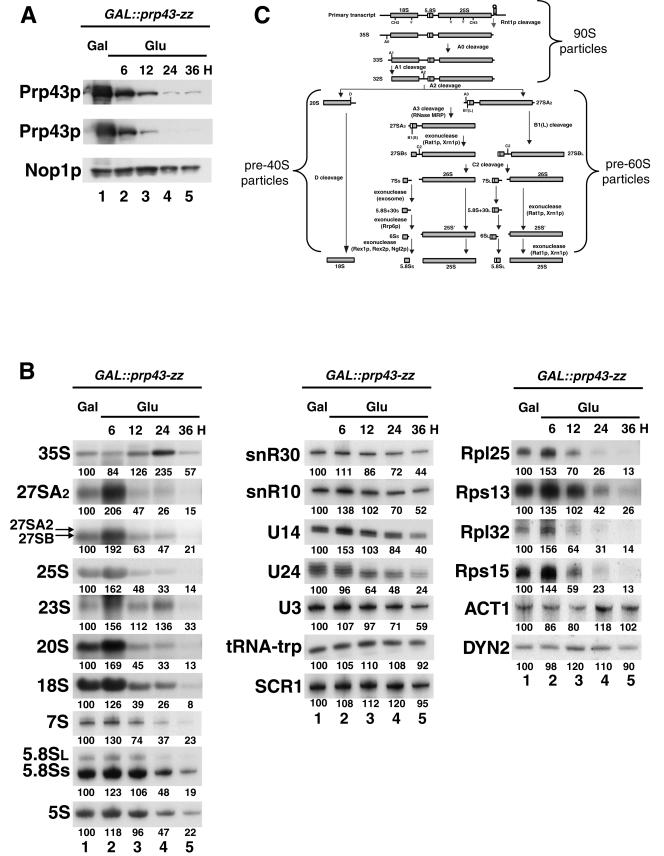
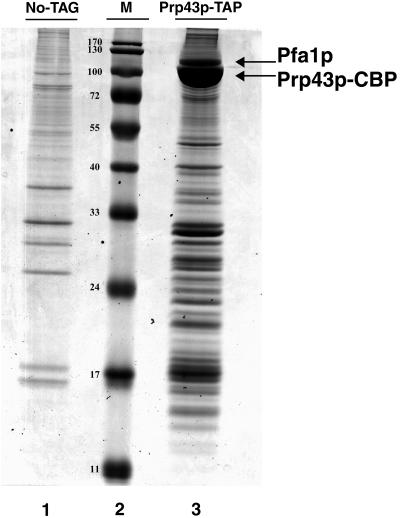
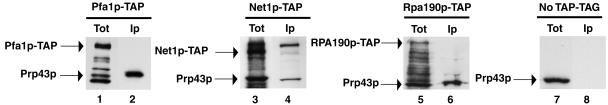
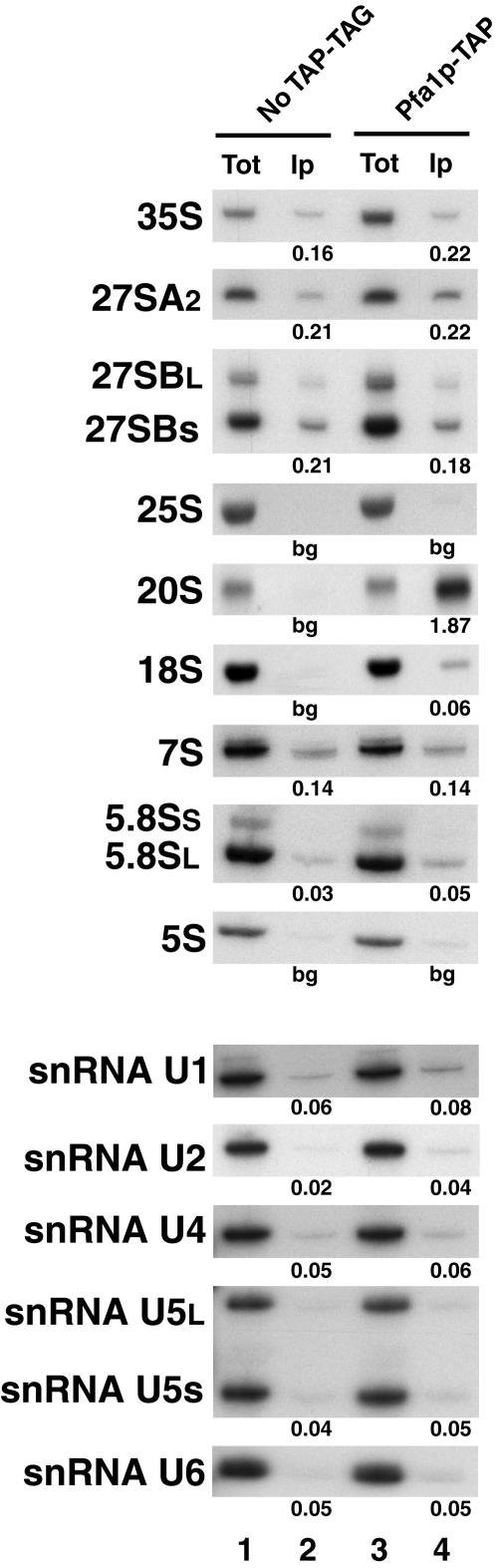
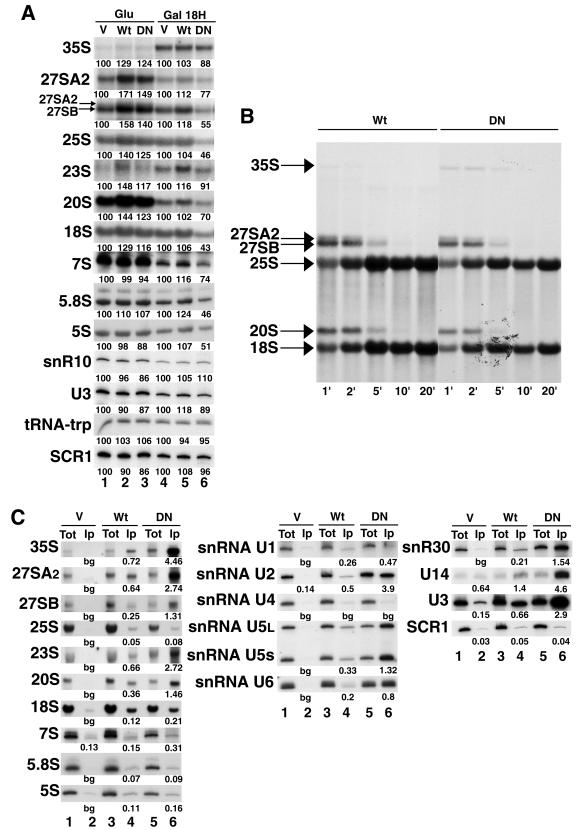
Prp43p is a putative helicase of the DEAH family which is required for the release of the lariat intron from the spliceosome. Prp43p could also play a role in ribosome synthesis, since it accumulates in the nucleolus. Consistent with this hypothesis, we find that depletion of Prp43p leads to accumulation of 35S pre-rRNA and strongly reduces levels of all downstream pre-rRNA processing intermediates. As a result, the steady-state levels of mature rRNAs are greatly diminished following Prp43p depletion. We present data arguing that such effects are unlikely to be solely due to splicing defects. Moreover, we demonstrate by a combination of a comprehensive two-hybrid screen, tandem-affinity purification followed by mass spectrometry, and Northern analyses that Prp43p is associated with 90S, pre-60S, and pre-40S ribosomal particles. Prp43p seems preferentially associated with Pfa1p, a novel specific component of pre-40S ribosomal particles. In addition, Prp43p interacts with components of the RNA polymerase I (Pol I) transcription machinery and with mature 18S and 25S rRNAs. Hence, Prp43p might be delivered to nascent 90S ribosomal particles during pre-rRNA transcription and remain associated with preribosomal particles until their final maturation steps in the cytoplasm. Our data also suggest that the ATPase activity of Prp43p is required for early steps of pre-rRNA processing and normal accumulation of mature rRNAs.
Figures

References
-
- Bassler, J., P. Grandi, O. Gadal, T. Lessmann, E. Petfalski, D. Tollervey, J. Lechner, and E. Hurt. 2001. Identification of a 60S preribosomal particle that is closely linked to nuclear export. Mol. Cell 8:517-529. - PubMed
-
- Becam, A. M., F. Nasr, W. J. Racki, M. Zagulski, and C. J. Herbert. 2001. Ria1p (Ynl163c), a protein similar to elongation factors 2, is involved in the biogenesis of the 60S subunit of the ribosome in Saccharomyces cerevisiae. Mol. Genet. Genom. 266:454-462. - PubMed
-
- Bonneaud, N., O. Ozier-Kalogeropoulos, G. Y. Li, M. Labouesse, L. Minvielle-Sebastia, and F. Lacroute. 1991. A family of low and high copy replicative, integrative and single-stranded S. cerevisiae/E. coli shuttle vectors. Yeast 7:609-615. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials