

Met/Hepatocyte growth factor receptor ubiquitination suppresses transformation and is required for Hrs phosphorylation
- PMID: 16227611
- PMCID: PMC1265818
- DOI: 10.1128/MCB.25.21.9632-9645.2005
Met/Hepatocyte growth factor receptor ubiquitination suppresses transformation and is required for Hrs phosphorylation
Abstract
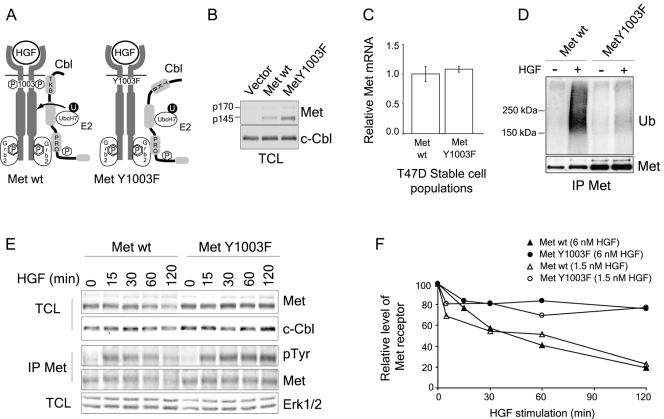
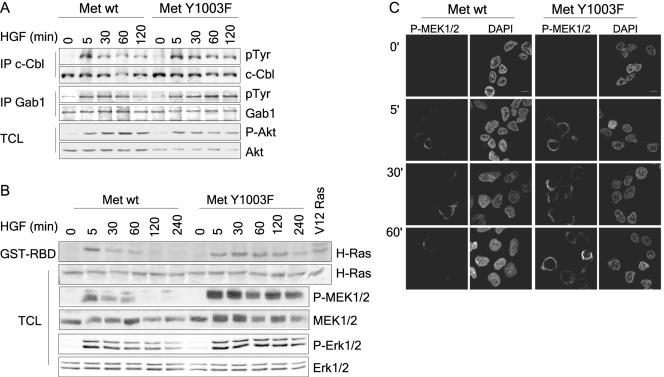
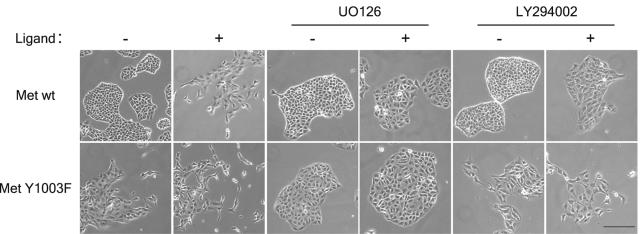
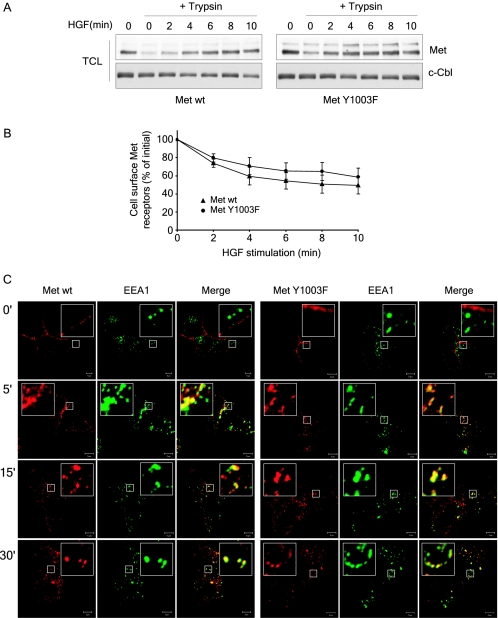
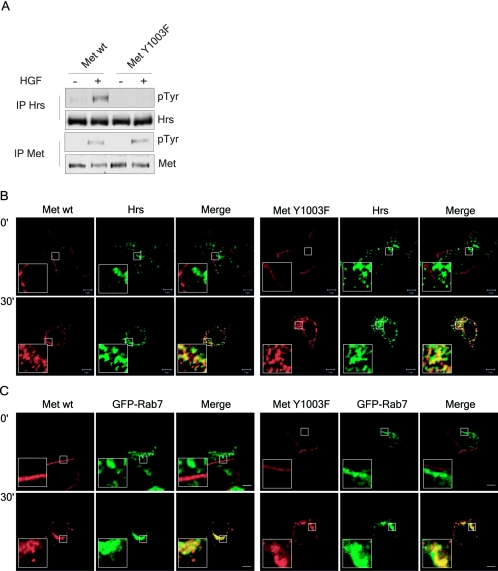
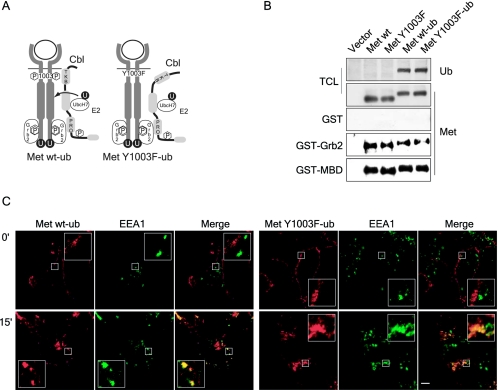
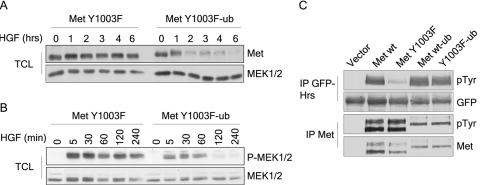
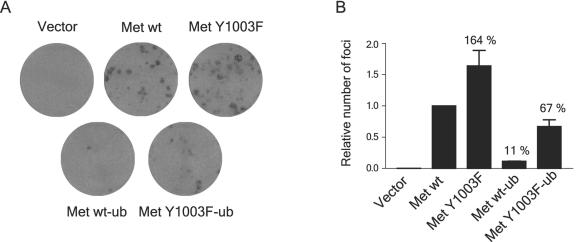
The Met receptor tyrosine kinase (RTK) regulates epithelial remodeling, dispersal, and invasion and is deregulated in many human cancers. It is now accepted that impaired down-regulation, as well as sustained activation, of RTKs could contribute to their deregulation. Down-regulation of the Met receptor involves ligand-induced internalization, ubiquitination by Cbl ubiquitin ligases, and lysosomal degradation. Here we report that a ubiquitination-deficient Met receptor mutant (Y1003F) is tumorigenic in vivo. The Met Y1003F mutant is internalized, and undergoes endosomal trafficking with kinetics similar to the wild-type Met receptor, yet is inefficiently targeted for degradation. This results in sustained activation of Met Y1003F and downstream signals involving the Ras-mitogen-activated protein kinase pathway, cell transformation, and tumorigenesis. Although Met Y1003F undergoes endosomal trafficking and localizes with the cargo-sorting protein Hrs, it is unable to induce phosphorylation of Hrs. Fusion of monoubiquitin to Met Y1003F is sufficient to decrease Met receptor stability and prevent sustained MEK1/2 activation. In addition, this rescues Hrs tyrosine phosphorylation and decreases transformation in a focus-forming assay. These results demonstrate that Cbl-dependent ubiquitination is dispensable for Met internalization but is critical to target the Met receptor to components of the lysosomal sorting machinery and to suppress its inherent transforming activity.
Figures

References
-
- Bache, K. G., C. Raiborg, A. Mehlum, and H. Stenmark. 2003. STAM and Hrs are subunits of a multivalent ubiquitin-binding complex on early endosomes. J. Biol. Chem. 278:12513-12521. - PubMed
-
- Bilodeau, P. S., J. L. Urbanowski, S. C. Winistorfer, and R. C. Piper. 2002. The Vps27p Hse1p complex binds ubiquitin and mediates endosomal protein sorting. Nat. Cell Biol. 4:534-539. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous