

Review
doi: 10.1038/nrm1745.
FtsZ and the division of prokaryotic cells and organelles
Affiliations
- PMID: 16227976
- PMCID: PMC4757588
- DOI: 10.1038/nrm1745
Item in Clipboard
Review
FtsZ and the division of prokaryotic cells and organelles
Nat Rev Mol Cell Biol.
2005 Nov.
Abstract
Binary fission of many prokaryotes as well as some eukaryotic organelles depends on the FtsZ protein, which self-assembles into a membrane-associated ring structure early in the division process. FtsZ is homologous to tubulin, the building block of the microtubule cytoskeleton in eukaryotes. Recent advances in genomics and cell-imaging techniques have paved the way for the remarkable progress in our understanding of fission in bacteria and organelles.
Conflict of interest statement
Competing interests statement
The author declares no competing financial interests.
Figures
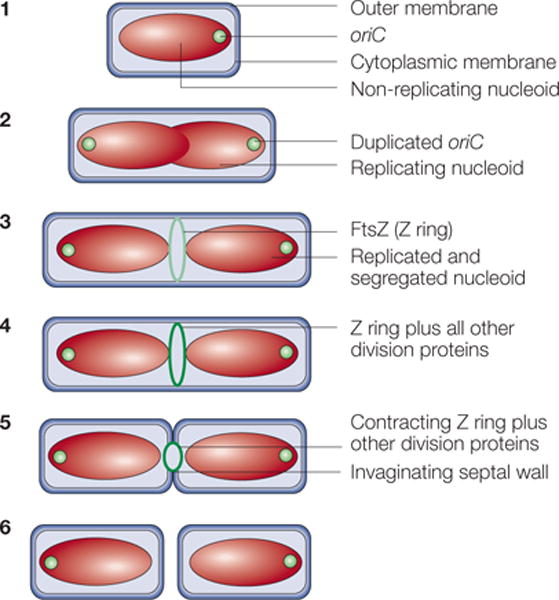
In step 1, newborn cells grown at low growth rates contain a single, non-replicating chromosome in a structure known as a nucleoid. Soon after chromosome replication initiates, the replication origins (oriC) move towards the cell poles until both daughter chromosomes are segregated (step 2). Near the end of this process, the FtsZ protein assembles into the Z ring on the inner face of the cytoplasmic membrane (light green ring) at the cell centre, marking the future division site (step 3). In step 4, the Z ring recruits at least ten membrane-associated proteins (for details, see FIG. 6) to assemble the cell-division protein machinery (dark green ring). This machinery synthesizes the division septum, which consists of cell-wall material, with the Z ring at the leading edge of membrane invagination. Contraction of the Z ring and constriction of the outer membrane follow (step 5). The result is the production of two separate newborn daughter cells (step 6).

The domain structure applies to most FtsZ proteins. The N terminus and variable spacer domains are highly variable in length, and their precise functions are unknown. The core region displays most similarity to tubulin and is required for GTP binding and hydrolysis as well as assembly into protofilaments. The C-terminal peptide interacts with other cell-division proteins recruited by FtsZ such as ZipA, FtsA and FtsW, and might function mainly to anchor the Z ring to the membrane using these proteins.
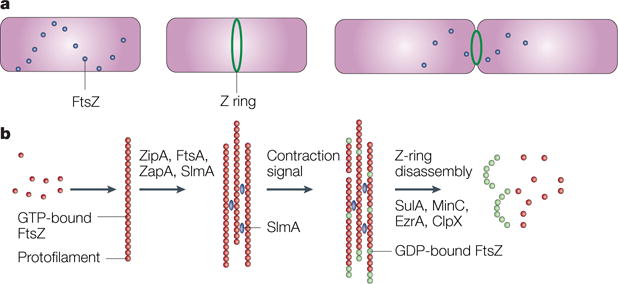
a | A time course, starting with a newborn Escherichia coli cell without a Z ring but with FtsZ in spiral patterns, followed by a cell with a Z ring, and finally, a dividing cell with a contracting Z ring. b | The putative time course of FtsZ assembly. Red dots indicate GTP-bound FtsZ dimers that assemble into protofilaments. GTP hydrolysis during assembly is probably balanced by rapid exchange of the GDP for the abundant GTP within the cell. Several protein factors, including ZipA, ZapA and probably also FtsA and the nucleoid occlusion protein SlmA (blue), bundle these protofilaments and anchor them to the cytoplasmic membrane. An unknown signal triggers ring contraction and probable disassembly; this might involve stimulation of GTP hydrolysis, which would increase the number of GDP-bound subunits at protofilament ends (shown in green), causing curved protofilaments to be formed. The free FtsZ then forms a spiral pattern throughout the cell, possibly by reassembling, and this FtsZ would then be available to form a new ring in the daughter cells. The presence of high levels of inhibitor proteins such as SulA, MinC, ClpX or EzrA (in Bacillus subtilis) might help to antagonize the ring-stabilizing proteins and tip the balance towards disassembly, although none of the inhibitor proteins is required for Z-ring contraction or disassembly.

a | A time course of a group of E. coli cells expressing FtsZ–GFP, showing the Z ring at the midcell but also displaying the oscillation of FtsZ from one side of the ring to the other. b | A schematic time course of the spatial regulation of FtsZ. The top four cells span approximately 2 minutes of growth of a cell early in the cell cycle, showing how the oscillation of Min proteins (pink) and FtsZ (green), along with nucleoid occlusion (yellow), prevent assembly of the Z ring (dark green) at all locations in the cell. The fifth cell is at a later point in the cell cycle, after chromosomal replication and segregation is well underway, relieving nucleoid occlusion at the cell centre. This, along with potential positive regulators, lowers the barrier for Z-ring assembly in a zone near the middle of the cell (green), allowing Z-ring assembly (final cell). Blue dots represent higher concentrations of non-ring FtsZ that oscillate in spiral patterns. Similar mechanisms might help to centre the Z ring in chloroplasts and in other bacteria that contain the Min system, such as cyanobacteria. Nucleoid occlusion might be the main spatial regulator in species that lack Min proteins.
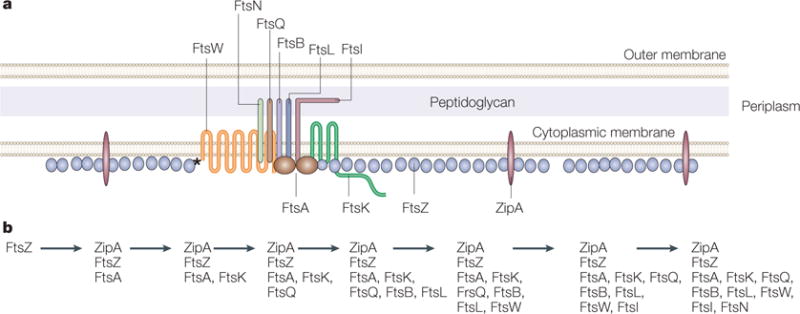
a | A model of the Escherichia coli Z ring and its essential protein partners is shown in cross section. FtsZ is shown as a series of single protofilaments at the membrane, although the actual structure of FtsZ in the Z ring is unknown. Both ZipA and FtsA contact FtsZ as well as the membrane in E. coli. However, FtsZ contacts FtsW directly in Mycobacterium tuberculosis (asterisk), which lacks ZipA and FtsA. A single transmembrane subassembly associated with an FtsA dimer is shown, based on the low relative amounts of most of the integral membrane proteins that are essential for cell division. These membrane proteins include FtsQ, FtsB, FtsL, FtsI and FtsN, which are bitopic proteins that each have a single transmembrane and periplasmic domain, and FtsW and FtsK, which are polytopic proteins with multiple transmembrane and periplasmic domains. The network of protein–protein associations is implied by the proximity of the proteins in the diagram. Proteins implicated in stabilization of the ring structure are labelled below the cytoplasmic membrane lines, whereas proteins implicated in later functions in septum formation, such as septum synthesis, are labelled above the lines. b | The dependency order of recruitment of essential cell-division proteins to the Z ring, as deduced from the requirement of a given protein for another’s localization to the Z ring.
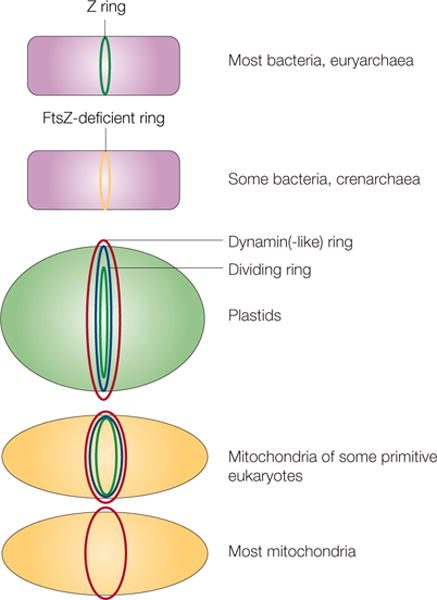
Different types of cells or organelles and their use of FtsZ or alternative proteins for fission are shown. In euryarchaea and many bacteria, FtsZ (green) localizes to the inner side of the inner membrane as the Z ring. Crenarchaea and some bacteria lack FtsZ, so some other protein must localize as a ring at the site of division (yellow). In plastids and mitochondria, dynamin or dynamin-like protein rings (red) localize to the cytoplasmic face. These organelles also contain a dividing ring (blue) and, in the case of chloroplasts and some primitive mitochondria, an innermost ring of FtsZ (green). Most mitochondria, including those of fungi and animals, lack FtsZ and a detectable dividing ring, but still rely on dynamin on the cytosolic face for fission (red).
References
-
- Bi E, Lutkenhaus J. FtsZ ring structure associated with division in Escherichia coli. Nature. 1991;354:161–164. - PubMed
-
- Weiss DS. Bacterial cell division and the septal ring. Mol Microbiol. 2004;54:588–597. - PubMed
-
- Vaughan S, Wickstead B, Gull K, Addinall SG. Molecular evolution of FtsZ protein sequences encoded within the genomes of archaea, bacteria, and eukaryota. J Mol Evol. 2004;58:19–29. - PubMed
-
- Sontag CA, Staley JT, Erickson HP. In vitro assembly and GTP hydrolysis by bacterial tubulins BtubA and BtubB. J Cell Biol. 2005;169:233–238. A member of the Chlamydia/Verrucomicrobia group of bacteria that lacks ftsZ contains instead two genes that are evolutionarily closer to tubulin than FtsZ. This paper investigates the assembly and nucleotide-binding properties of these two bacterial tubulins. - PMC - PubMed
-
- Glass JI, et al. The complete sequence of the mucosal pathogen Ureaplasma urealyticum. Nature. 2000;407:757–762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
