

Lineage-specific variations of congruent evolution among DNA sequences from three genomes, and relaxed selective constraints on rbcL in Cryptomonas (Cryptophyceae)
- PMID: 16232313
- PMCID: PMC1285359
- DOI: 10.1186/1471-2148-5-56
Lineage-specific variations of congruent evolution among DNA sequences from three genomes, and relaxed selective constraints on rbcL in Cryptomonas (Cryptophyceae)
Abstract
Background: Plastid-bearing cryptophytes like Cryptomonas contain four genomes in a cell, the nucleus, the nucleomorph, the plastid genome and the mitochondrial genome. Comparative phylogenetic analyses encompassing DNA sequences from three different genomes were performed on nineteen photosynthetic and four colorless Cryptomonas strains. Twenty-three rbcL genes and fourteen nuclear SSU rDNA sequences were newly sequenced to examine the impact of photosynthesis loss on codon usage in the rbcL genes, and to compare the rbcL gene phylogeny in terms of tree topology and evolutionary rates with phylogenies inferred from nuclear ribosomal DNA (concatenated SSU rDNA, ITS2 and partial LSU rDNA), and nucleomorph SSU rDNA.
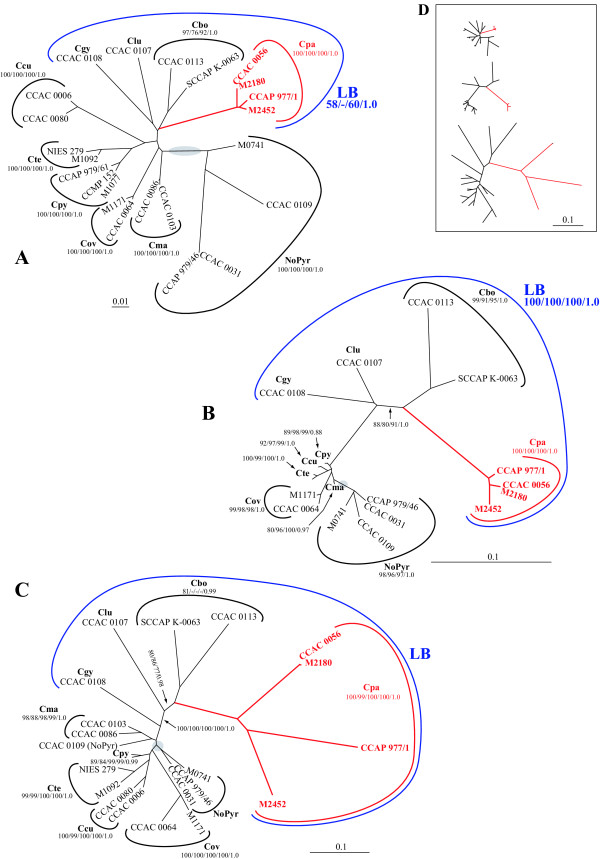
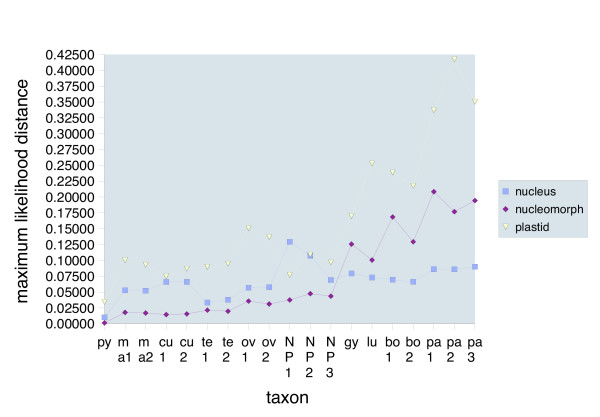
Results: Largely congruent branching patterns and accelerated evolutionary rates were found in nucleomorph SSU rDNA and rbcL genes in a clade that consisted of photosynthetic and colorless species suggesting a coevolution of the two genomes. The extremely accelerated rates in the rbcL phylogeny correlated with a shift from selection to mutation drift in codon usage of two-fold degenerate NNY codons comprising the amino acids asparagine, aspartate, histidine, phenylalanine, and tyrosine. Cysteine was the sole exception. The shift in codon usage seemed to follow a gradient from early diverging photosynthetic to late diverging photosynthetic or heterotrophic taxa along the branches. In the early branching taxa, codon preferences were changed in one to two amino acids, whereas in the late diverging taxa, including the colorless strains, between four and five amino acids showed changes in codon usage.
Conclusion: Nucleomorph and plastid gene phylogenies indicate that loss of photosynthesis in the colorless Cryptomonas strains examined in this study possibly was the result of accelerated evolutionary rates that started already in photosynthetic ancestors. Shifts in codon usage are usually considered to be caused by changes in functional constraints and in gene expression levels. Thus, the increasing influence of mutation drift on codon usage along the clade may indicate gradually relaxed constraints and reduced expression levels on the rbcL gene, finally correlating with a loss of photosynthesis in the colorless Cryptomonas paramaecium strains.
Figures
Similar articles
-
Gene loss, pseudogenization, and independent genome reduction in non-photosynthetic species of Cryptomonas (Cryptophyceae) revealed by comparative nucleomorph genomics.BMC Biol. 2022 Oct 8;20(1):227. doi: 10.1186/s12915-022-01429-6. BMC Biol. 2022. PMID: 36209116 Free PMC article.
-
Comparative Plastid Genomics of Cryptomonas Species Reveals Fine-Scale Genomic Responses to Loss of Photosynthesis.Genome Biol Evol. 2020 Feb 1;12(2):3926-3937. doi: 10.1093/gbe/evaa001. Genome Biol Evol. 2020. PMID: 31922581 Free PMC article.
-
Multiple independent losses of photosynthesis and differing evolutionary rates in the genus Cryptomonas (Cryptophyceae): combined phylogenetic analyses of DNA sequences of the nuclear and the nucleomorph ribosomal operons.J Mol Evol. 2005 Feb;60(2):183-95. doi: 10.1007/s00239-004-0089-5. J Mol Evol. 2005. PMID: 15785847
-
Ubiquitin fusion proteins in algae: implications for cell biology and the spread of photosynthesis.BMC Genomics. 2019 Jan 14;20(1):38. doi: 10.1186/s12864-018-5412-4. BMC Genomics. 2019. PMID: 30642248 Free PMC article. Review.
-
Rates and patterns of chloroplast DNA evolution.Proc Natl Acad Sci U S A. 1994 Jul 19;91(15):6795-801. doi: 10.1073/pnas.91.15.6795. Proc Natl Acad Sci U S A. 1994. PMID: 8041699 Free PMC article. Review.
Cited by
-
The complete plastid genome sequence of the secondarily nonphotosynthetic alga Cryptomonas paramecium: reduction, compaction, and accelerated evolutionary rate.Genome Biol Evol. 2009 Nov 13;1:439-48. doi: 10.1093/gbe/evp047. Genome Biol Evol. 2009. PMID: 20333213 Free PMC article.
-
The Genetic Diversity of Mesodinium and Associated Cryptophytes.Front Microbiol. 2016 Dec 20;7:2017. doi: 10.3389/fmicb.2016.02017. eCollection 2016. Front Microbiol. 2016. PMID: 28066344 Free PMC article.
-
Gene loss, pseudogenization, and independent genome reduction in non-photosynthetic species of Cryptomonas (Cryptophyceae) revealed by comparative nucleomorph genomics.BMC Biol. 2022 Oct 8;20(1):227. doi: 10.1186/s12915-022-01429-6. BMC Biol. 2022. PMID: 36209116 Free PMC article.
-
Evolution of heterotrophy in chrysophytes as reflected by comparative transcriptomics.FEMS Microbiol Ecol. 2018 Apr 1;94(4):fiy039. doi: 10.1093/femsec/fiy039. FEMS Microbiol Ecol. 2018. PMID: 29518196 Free PMC article.
-
Comparative Plastid Genomics of Cryptomonas Species Reveals Fine-Scale Genomic Responses to Loss of Photosynthesis.Genome Biol Evol. 2020 Feb 1;12(2):3926-3937. doi: 10.1093/gbe/evaa001. Genome Biol Evol. 2020. PMID: 31922581 Free PMC article.
References
-
- Kellogg EA, Juliano ND. The structure and function of RuBisCO and their implications for systematic studies. Am J Bot. 1997;84:413–428. - PubMed
-
- Tabita FR. Microbial ribulose 1,5-bisphosphate carboxylase/oxygenase: A different perspective. Photosynthesis Res. 1999;60:1–28. doi: 10.1023/A:1006211417981. - DOI
-
- Rodermel S. Subunit control of Rubisco biosynthesis – a relic of an endosymbiotic past? Photosynthesis Res. 1999;59:105–123. doi: 10.1023/A:1006122619851. - DOI
-
- Ellis RJ. The most abundant protein in the world. Trends Biochem Sci. 1979;4:241–244. doi: 10.1016/0968-0004(79)90212-3. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
