

Contribution of the C-terminal tri-lysine regions of human immunodeficiency virus type 1 integrase for efficient reverse transcription and viral DNA nuclear import
- PMID: 16232319
- PMCID: PMC1277849
- DOI: 10.1186/1742-4690-2-62
Contribution of the C-terminal tri-lysine regions of human immunodeficiency virus type 1 integrase for efficient reverse transcription and viral DNA nuclear import
Abstract
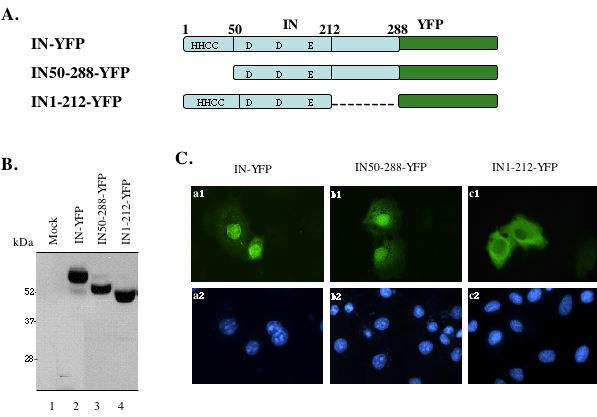
Background: In addition to mediating the integration process, HIV-1 integrase (IN) has also been implicated in different steps during viral life cycle including reverse transcription and viral DNA nuclear import. Although the karyophilic property of HIV-1 IN has been well demonstrated using a variety of experimental approaches, the definition of domain(s) and/or motif(s) within the protein that mediate viral DNA nuclear import and its mechanism are still disputed and controversial. In this study, we performed mutagenic analyses to investigate the contribution of different regions in the C-terminal domain of HIV-1 IN to protein nuclear localization as well as their effects on virus infection.
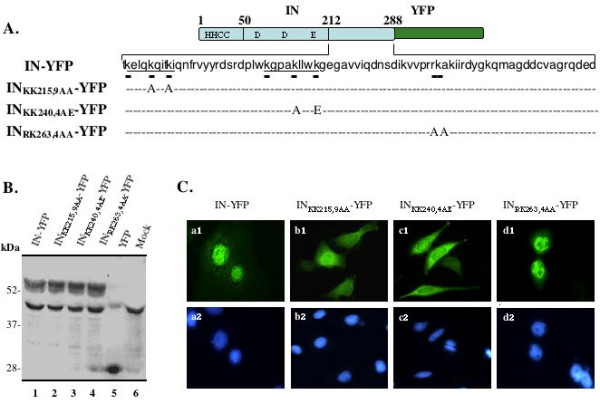
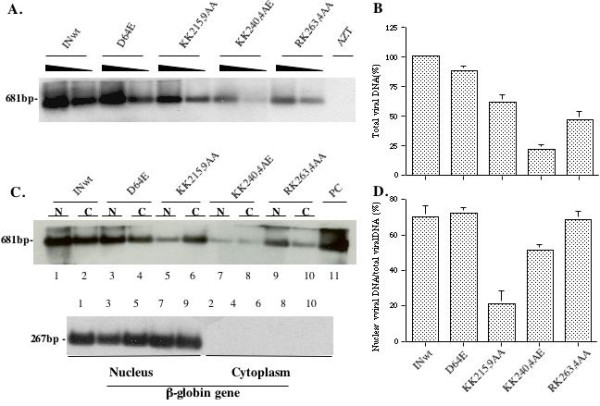
Results: Our analysis showed that replacing lysine residues in two highly conserved tri-lysine regions, which are located within previously described Region C (235WKGPAKLLWKGEGAVV) and sequence Q (211KELQKQITK) in the C-terminal domain of HIV-1 IN, impaired protein nuclear accumulation, while mutations for RK263,4 had no significant effect. Analysis of their effects on viral infection in a VSV-G pseudotyped RT/IN trans-complemented HIV-1 single cycle replication system revealed that all three C-terminal mutant viruses (KK215,9AA, KK240,4AE and RK263,4AA) exhibited more severe defect of induction of beta-Gal positive cells and luciferase activity than an IN class 1 mutant D64E in HeLa-CD4-CCR5-beta-Gal cells, and in dividing as well as non-dividing C8166 T cells, suggesting that some viral defects are occurring prior to viral integration. Furthermore, by analyzing viral DNA synthesis and the nucleus-associated viral DNA level, the results clearly showed that, although all three C-terminal mutants inhibited viral reverse transcription to different extents, the KK240,4AE mutant exhibited most profound effect on this step, whereas KK215,9AA significantly impaired viral DNA nuclear import. In addition, our analysis could not detect viral DNA integration in each C-terminal mutant infection, even though they displayed various low levels of nucleus-associated viral DNA, suggesting that these C-terminal mutants also impaired viral DNA integration ability.
Conclusion: All of these results indicate that, in addition to being involved in HIV-1 reverse transcription and integration, the C-terminal tri-lysine regions of IN also contribute to efficient viral DNA nuclear import during the early stage of HIV-1 replication.
Figures



References
-
- Ikeda T, Nishitsuji H, Zhou X, Nara N, Ohashi T, Kannagi M, Masuda T. Evaluation of the functional involvement of human immunodeficiency virus type 1 integrase in nuclear import of viral cDNA during acute infection. J Virol. 2004;78:11563–11573. doi: 10.1128/JVI.78.21.11563-11573.2004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
