

Mice deficient in Ext2 lack heparan sulfate and develop exostoses
- PMID: 16236767
- PMCID: PMC2767329
- DOI: 10.1242/dev.02088
Mice deficient in Ext2 lack heparan sulfate and develop exostoses
Abstract
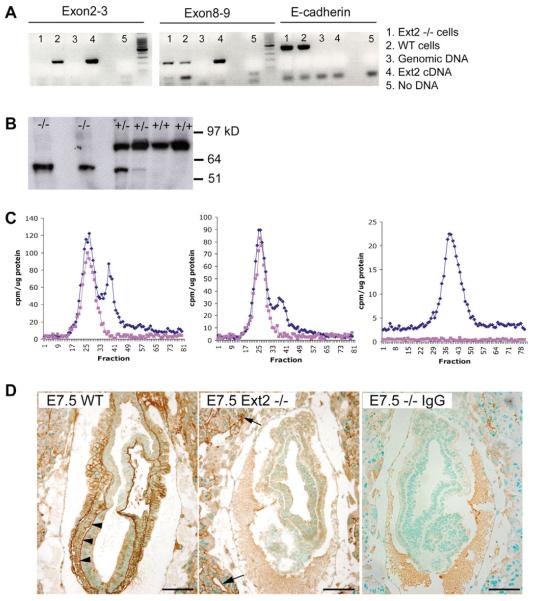
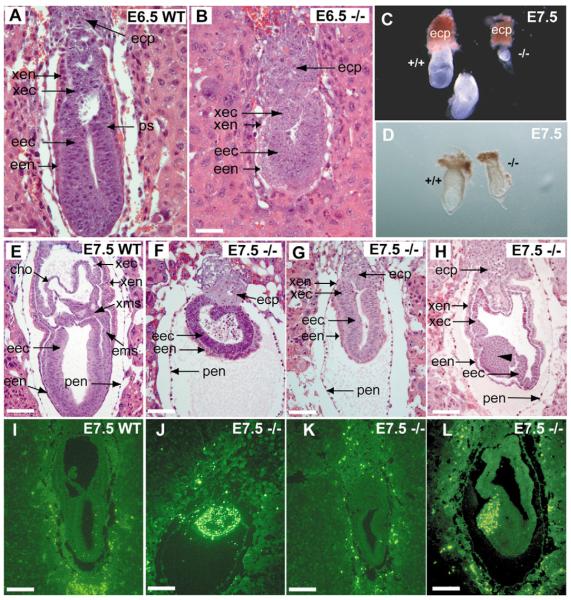
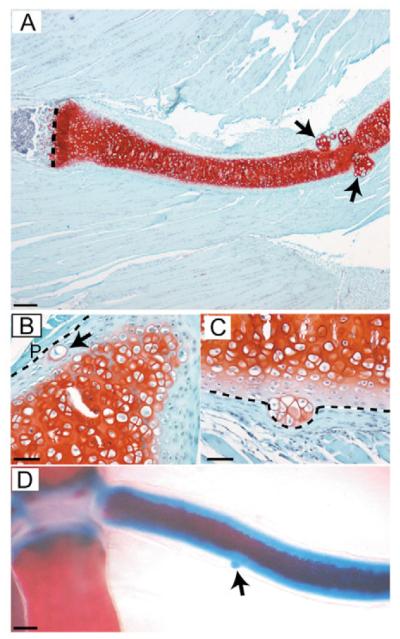
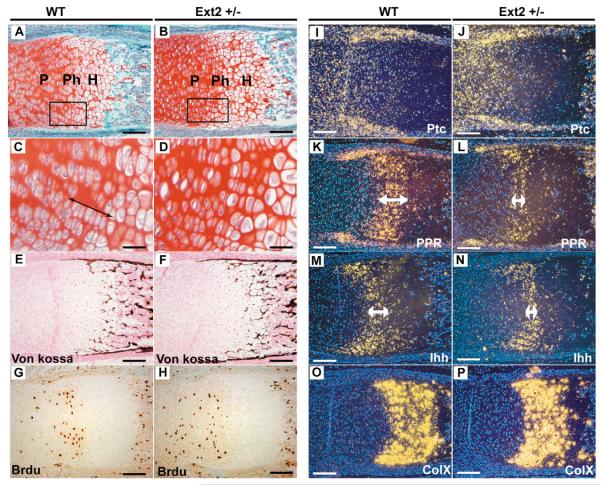
Hereditary multiple exostoses (HME) is a genetically heterogeneous human disease characterized by the development of bony outgrowths near the ends of long bones. HME results from mutations in EXT1 and EXT2, genes that encode glycosyltransferases that synthesize heparan sulfate chains. To study the relationship of the disease to mutations in these genes, we generated Ext2-null mice by gene targeting. Homozygous mutant embryos developed normally until embryonic day 6.0, when they became growth arrested and failed to gastrulate, pointing to the early essential role for heparan sulfate in developing embryos. Heterozygotes had a normal lifespan and were fertile; however, analysis of their skeletons showed that about one-third of the animals formed one or more ectopic bone growths (exostoses). Significantly, all of the mice showed multiple abnormalities in cartilage differentiation, including disorganization of chondrocytes in long bones and premature hypertrophy in costochondral cartilage. These changes were not attributable to a defect in hedgehog signaling, suggesting that they arise from deficiencies in other heparan sulfate-dependent pathways. The finding that haploinsufficiency triggers abnormal cartilage differentiation gives insight into the complex molecular mechanisms underlying the development of exostoses.
Figures





References
-
- Ahn J, Ludecke HJ, Lindow S, Horton WA, Lee B, Wagner MJ, Horsthemke B, Wells DE. Cloning of the putative tumour suppressor gene for hereditary multiple exostoses (EXT1) Nat. Genet. 1995;11:137–143. - PubMed
-
- Albrecht U, Eichele G, Helms JA, Lu H. Visualization of gene expression patterns by in situ hybridization. In: Daston G, editor. Molecular and Cellular Methods in Developmental Toxicology. CRC Press; Boca Raton: 1997. pp. 23–48.
-
- Ang SL, Conlon RA, Jin O, Rossant J. Positive and negative signals from mesoderm regulate the expression of mouse Otx2 in ectoderm explants. Development. 1994;120:2979–2989. - PubMed
-
- Bame KJ, Esko JD. Undersulfated heparan sulfate in a Chinese hamster ovary cell mutant defective in heparan sulfate N-sulfotransferase. J. Biol. Chem. 1989;264:8059–8065. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
