

K-ras4B and prenylated proteins lacking "second signals" associate dynamically with cellular membranes
- PMID: 16236799
- PMCID: PMC1345658
- DOI: 10.1091/mbc.e05-05-0408
K-ras4B and prenylated proteins lacking "second signals" associate dynamically with cellular membranes
Abstract
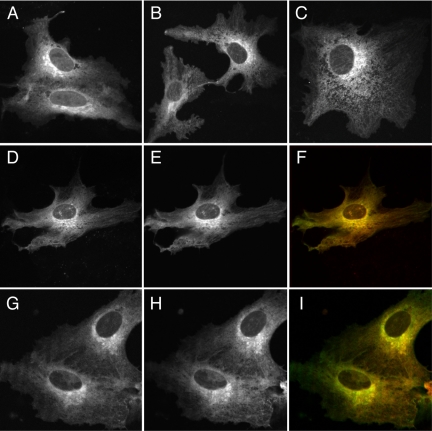
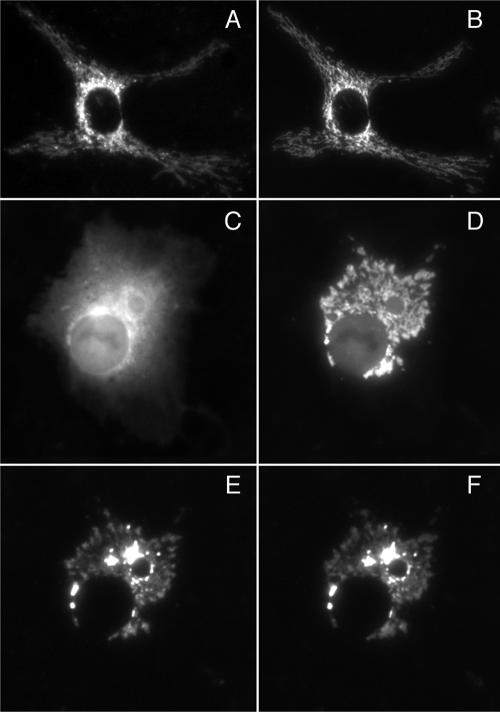
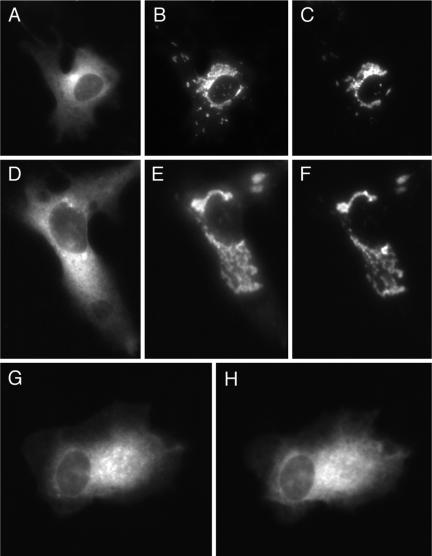
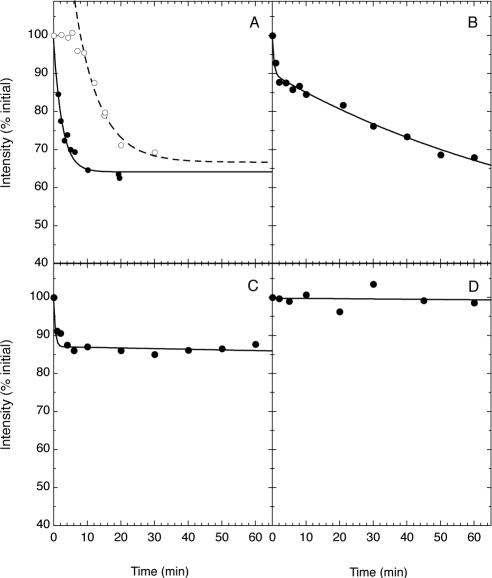
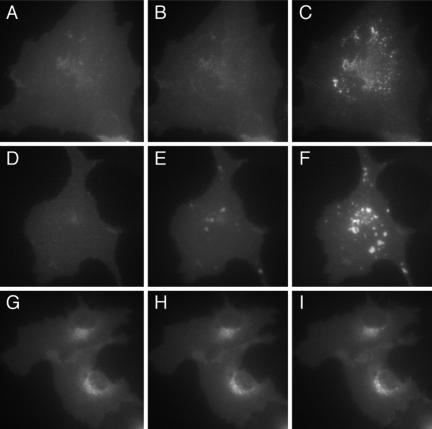
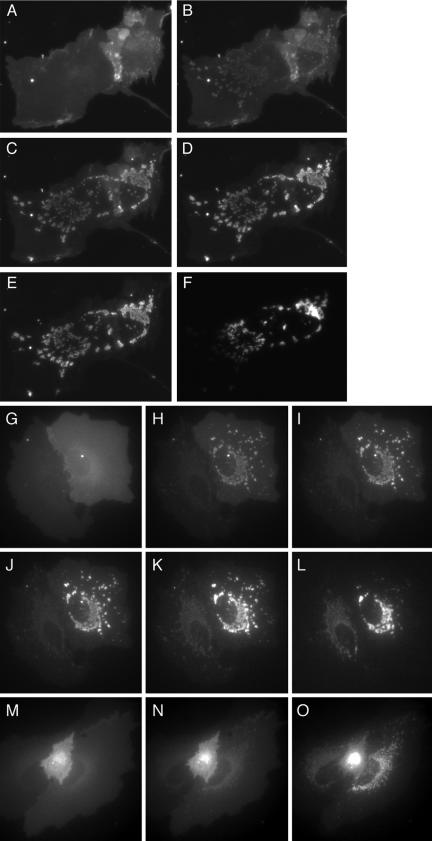
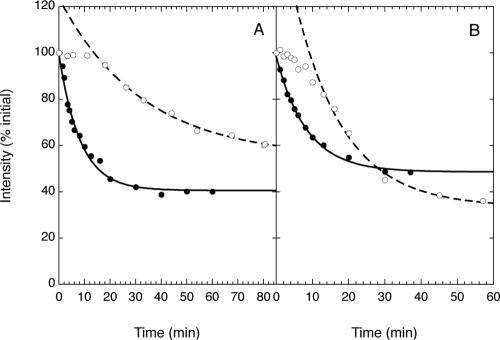
We have used fluorescence microscopy and the technique of rapamycin-regulated protein heterodimerization to examine the dynamics of the subcellular localizations of fluorescent proteins fused to lipid-modified protein sequences and to wild-type and mutated forms of full-length K-ras4B. Singly prenylated or myristoylated fluorescent protein derivatives lacking a "second signal" to direct them to specific subcellular destinations, but incorporating a rapamycin-dependent heterodimerization module, rapidly translocate to mitochondria upon rapamycin addition to bind to a mitochondrial outer membrane protein incorporating a complementary heterodimerization module. Under the same conditions analogous constructs anchored to the plasma membrane by multiply lipid-modified sequences, or by a transmembrane helix, show very slow or no transfer to mitochondria, respectively. Interestingly, however, fluorescent protein constructs incorporating either full-length K-ras4B or its plasma membrane-targeting sequence alone undergo rapamycin-induced transfer from the plasma membrane to mitochondria on a time scale of minutes, demonstrating the rapidly reversible nature of K-ras4B binding to the plasma membrane. The dynamic nature of the plasma membrane targeting of K-ras4B could contribute to K-ras4B function by facilitating redistribution of the protein between subcellular compartments under particular conditions.
Figures
References
-
- Baker, T. L., Booden, M. A., and Buss, J. E. (2000). S-Nitrosocysteine increases palmitate turnover on Ha-Ras in NIH 3T3 cells. J. Biol. Chem. 275, 22037–22047. - PubMed
-
- Baker, T. L., Zheng, H., Walker, J., Coloff, J. L., and Buss, J. E. (2003). Distinct rates of palmitate turnover on membrane-bound cellular and oncogenic Hras. J. Biol. Chem. 278, 19292–19300. - PubMed
-
- Bivona, T. G., Perez De Castro, I., Ahearn, I. M., Grana, T. M., Chiu, V. K., Lockyer, P. J., Cullen, P. J., Pellicer, A., Cox, A. D., and Philips, M. R. (2003). Phospholipase Cγ activates Ras on the Golgi apparatus by means of RasGRP1. Nature 424, 694–698. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
