

Chironomus tentans-repressor splicing factor represses SR protein function locally on pre-mRNA exons and is displaced at correct splice sites
- PMID: 16236800
- PMCID: PMC1345644
- DOI: 10.1091/mbc.e05-04-0339
Chironomus tentans-repressor splicing factor represses SR protein function locally on pre-mRNA exons and is displaced at correct splice sites
Abstract
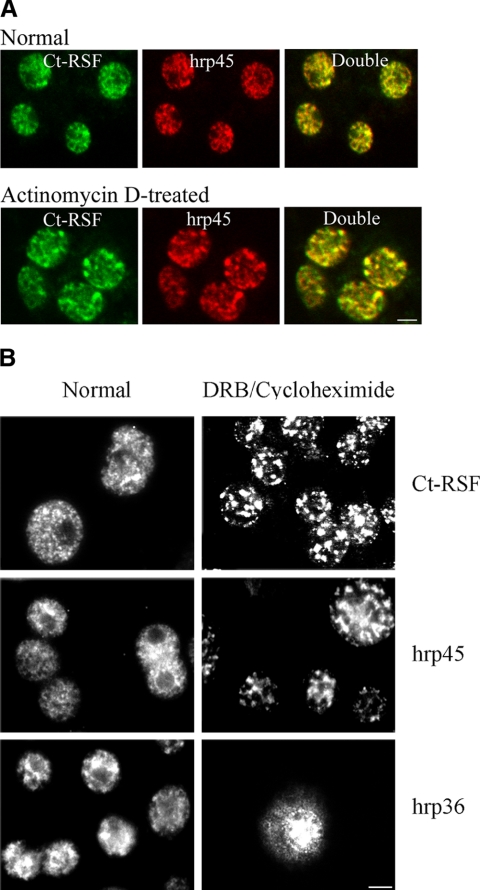
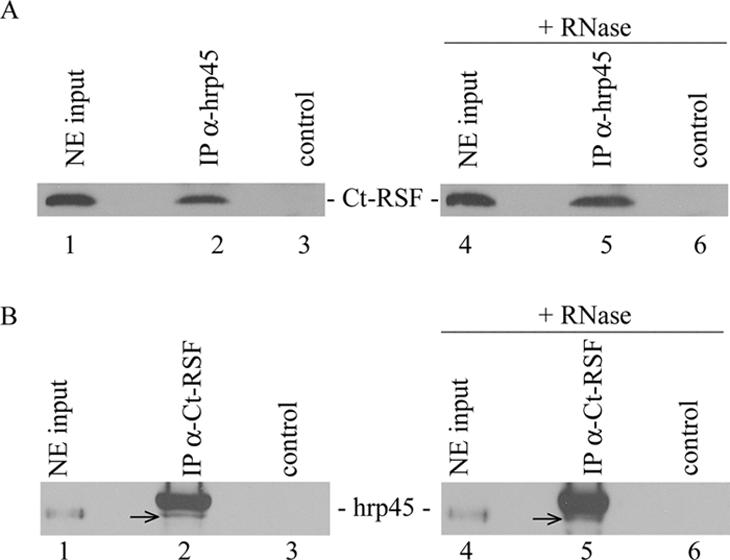
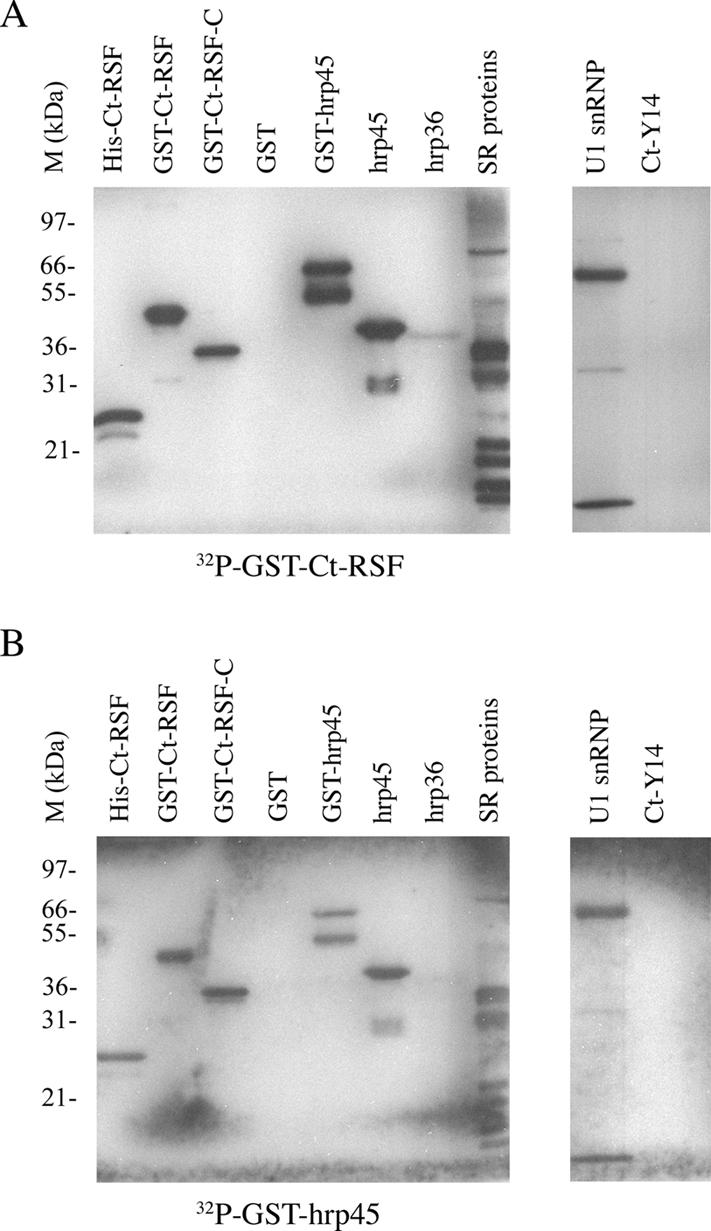
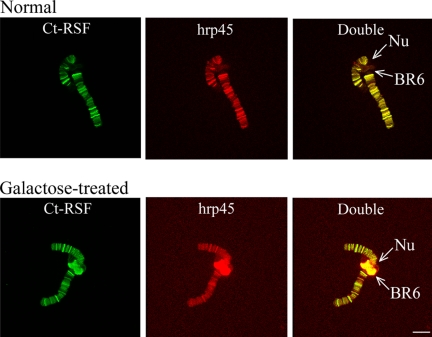
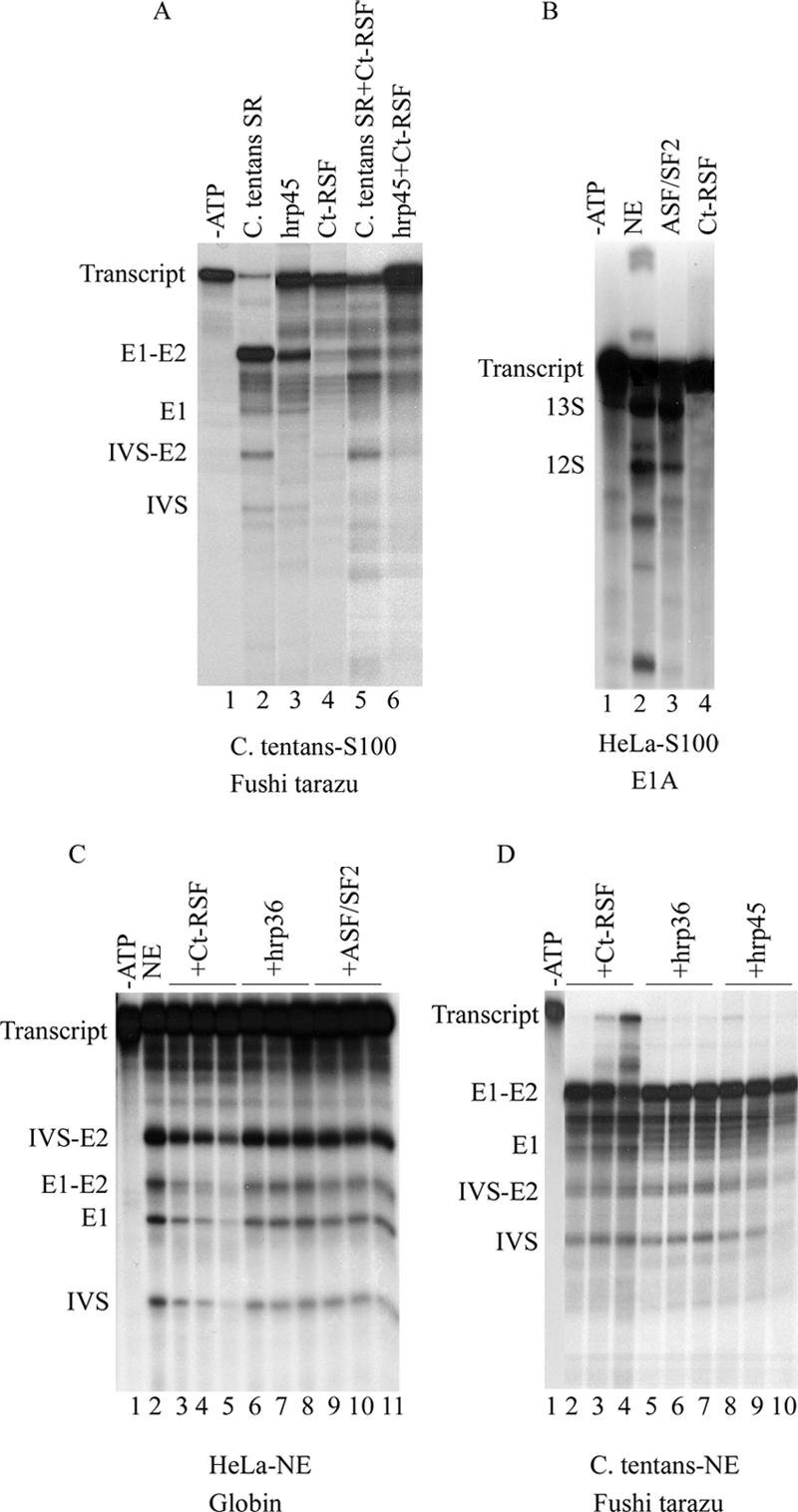
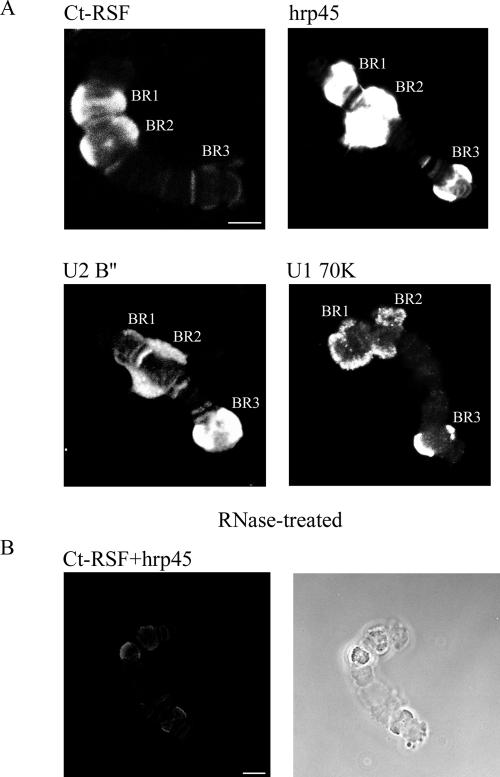
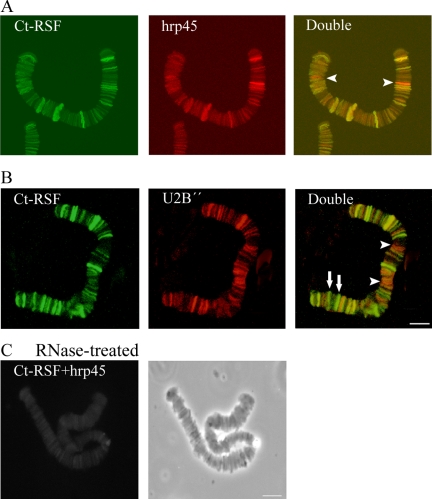
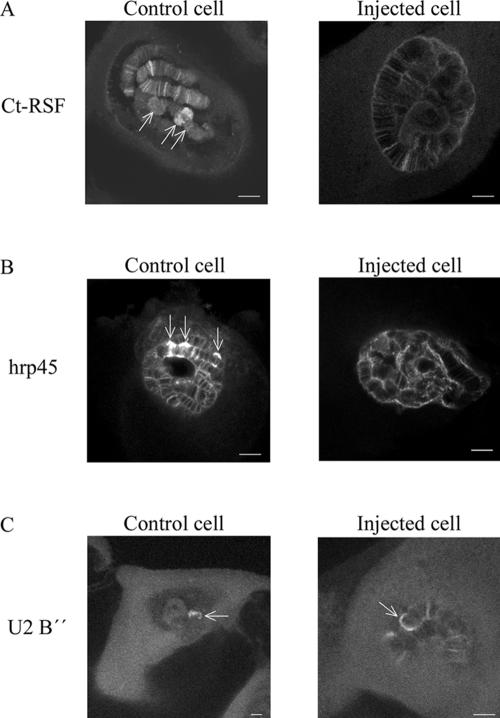
Chironomus tentans-repressor splicing factor (Ct-RSF) represses the activation of splicing by SR proteins in vitro. Ct-RSF colocalizes with the Ser-Arg-rich (SR) protein hrp45 in interchromatin granule clusters and coimmunoprecipitates with hrp45 in nuclear extracts. Ct-RSF and hrp45 can also interact directly in vitro. Ct-RSF and hrp45 are recruited together to transcribing genes and associate with growing pre-mRNAs. Ct-RSF and hrp45 colocalize at a large number of gene loci. Injection of anti-Ct-RSF antibodies into nuclei of living cells blocks association of both Ct-RSF and hrp45 with the growing pre-mRNA, whereas binding of U2 small nuclear ribonucleoprotein particle (snRNP) to the pre-mRNA is unaffected. On the intron-rich Balbiani ring (BR) 3 pre-mRNA, hrp45 as well as U1 and U2 snRNPs bind extensively, whereas relatively little Ct-RSF is present. In contrast, the BR1 and BR2 pre-mRNAs, dominated by exon sequences, bind relatively much Ct-RSF compared with hrp45 and snRNPs. Our data suggest that Ct-RSF represses SR protein function at exons and that the assembly of spliceosomes at authentic splice sites displaces Ct-RSF locally.
Figures

References
-
- Alzhanova-Ericsson, A. T., Sun, X., Visa, N., Kiseleva, E., Wurtz, T., and Daneholt, B. (1996). A protein of the SR family of splicing factors binds extensively to exonic Balbiani ring pre-mRNA and accompanies the RNA from the gene to the nuclear pore. Genes Dev. 10, 2881–2893. - PubMed
-
- Baurén, G., and Wieslander, L. (1994). Splicing of Balbiani ring 1 pre-mRNA occurs simultaneously with transcription. Cell 76, 183–192. - PubMed
-
- Bentley, D. (2002). The mRNA assembly line: transcription and processing machines in the same factory. Curr. Opin. Cell Biol. 14, 336 –342. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
