

Structure and increased thermostability of Rhodococcus sp. naphthalene 1,2-dioxygenase
- PMID: 16237006
- PMCID: PMC1272967
- DOI: 10.1128/JB.187.21.7222-7231.2005
Structure and increased thermostability of Rhodococcus sp. naphthalene 1,2-dioxygenase
Abstract
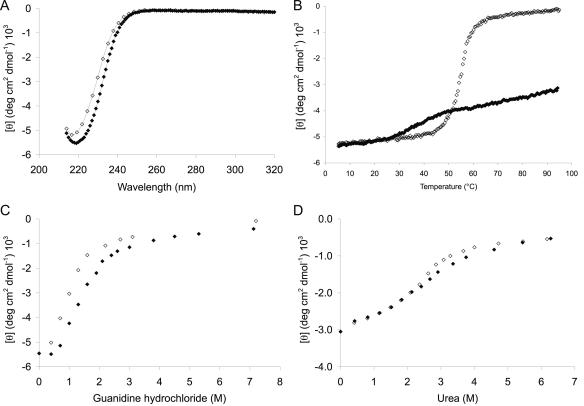
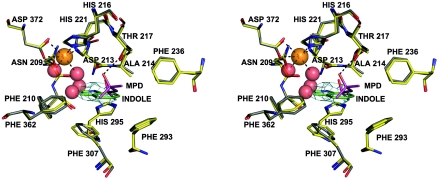
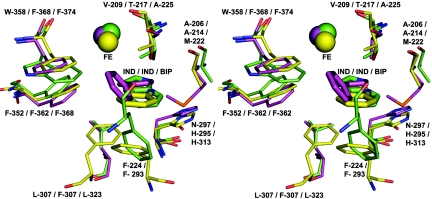
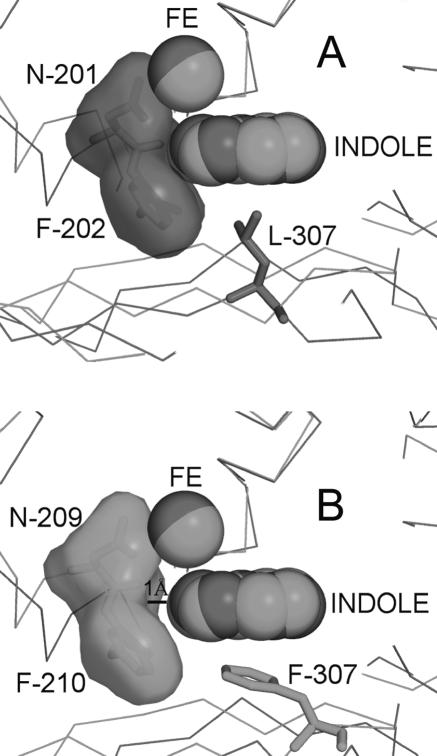
Rieske nonheme iron oxygenases form a large class of aromatic ring-hydroxylating dioxygenases found in microorganisms. These enzymes enable microorganisms to tolerate and even exclusively utilize aromatic compounds for growth, making them good candidates for use in synthesis of chiral intermediates and bioremediation. Studies of the chemical stability and thermostability of these enzymes thus become important. We report here the structure of free and substrate (indole)-bound forms of naphthalene dioxygenase from Rhodococcus sp. strain NCIMB12038. The structure of the Rhodococcus enzyme reveals that, despite a approximately 30% sequence identity between these naphthalene dioxygenases, their overall structures superpose very well with a root mean square deviation of less than 1.6 A. The differences in the active site of the two enzymes are pronounced near the entrance; however, indole binds to the Rhodococcus enzyme in the same orientation as in the Pseudomonas enzyme. Circular dichroism spectroscopy experiments show that the Rhodococcus enzyme has higher thermostability than the naphthalene dioxygenase from Pseudomonas species. The Pseudomonas enzyme has an apparent melting temperature of 55 degrees C while the Rhodococcus enzyme does not completely unfold even at 95 degrees C. Both enzymes, however, show similar unfolding behavior in urea, and the Rhodococcus enzyme is only slightly more tolerant to unfolding by guanidine hydrochloride. Structure analysis suggests that the higher thermostability of the Rhodococcus enzyme may be attributed to a larger buried surface area and extra salt bridge networks between the alpha and beta subunits in the Rhodococcus enzyme.
Figures

Similar articles
-
The role of active-site residues in naphthalene dioxygenase.J Ind Microbiol Biotechnol. 2003 May;30(5):271-8. doi: 10.1007/s10295-003-0043-3. Epub 2003 Apr 15. J Ind Microbiol Biotechnol. 2003. PMID: 12695887 Review.
-
Crystallization and preliminary X-ray diffraction analysis of naphthalene dioxygenase from Rhodococcus sp. strain NCIMB 12038.Acta Crystallogr D Biol Crystallogr. 2002 Dec;58(Pt 12):2173-4. doi: 10.1107/s0907444902016803. Epub 2002 Nov 23. Acta Crystallogr D Biol Crystallogr. 2002. PMID: 12454491
-
Purification and characterization of a novel naphthalene dioxygenase from Rhodococcus sp. strain NCIMB12038.J Bacteriol. 1999 Oct;181(19):6200-4. doi: 10.1128/JB.181.19.6200-6204.1999. J Bacteriol. 1999. PMID: 10498739 Free PMC article.
-
Substrate binding site of naphthalene 1,2-dioxygenase: functional implications of indole binding.J Mol Biol. 2000 Feb 18;296(2):701-12. doi: 10.1006/jmbi.1999.3462. J Mol Biol. 2000. PMID: 10669618
-
Engineering dioxygenases for efficient degradation of environmental pollutants.Curr Opin Biotechnol. 2000 Jun;11(3):244-9. doi: 10.1016/s0958-1669(00)00091-4. Curr Opin Biotechnol. 2000. PMID: 10851151 Review.
Cited by
-
Structural basis for divergent C-H hydroxylation selectivity in two Rieske oxygenases.Nat Commun. 2020 Jun 12;11(1):2991. doi: 10.1038/s41467-020-16729-0. Nat Commun. 2020. PMID: 32532989 Free PMC article.
-
Spectroscopic Characterisation of the Naphthalene Dioxygenase from Rhodococcus sp. Strain NCIMB12038.Int J Mol Sci. 2019 Jul 11;20(14):3402. doi: 10.3390/ijms20143402. Int J Mol Sci. 2019. PMID: 31373299 Free PMC article.
-
Characterization of a novel aromatic ring-hydroxylating oxygenase, NarA2B2, from thermophilic Hydrogenibacillus sp. strain N12.Appl Environ Microbiol. 2023 Oct 31;89(10):e0086523. doi: 10.1128/aem.00865-23. Epub 2023 Oct 11. Appl Environ Microbiol. 2023. PMID: 37819076 Free PMC article.
-
The Cell Shape-determining Csd6 Protein from Helicobacter pylori Constitutes a New Family of L,D-Carboxypeptidase.J Biol Chem. 2015 Oct 9;290(41):25103-17. doi: 10.1074/jbc.M115.658781. Epub 2015 Aug 25. J Biol Chem. 2015. PMID: 26306031 Free PMC article.
-
Near-IR MCD of the nonheme ferrous active site in naphthalene 1,2-dioxygenase: correlation to crystallography and structural insight into the mechanism of Rieske dioxygenases.J Am Chem Soc. 2008 Feb 6;130(5):1601-10. doi: 10.1021/ja074769o. Epub 2008 Jan 12. J Am Chem Soc. 2008. PMID: 18189388 Free PMC article.
References
-
- Arnone, M. I., L. Birolo, S. Pascarella, M. V. Cubellis, F. Bossa, G. Sannia, and G. Marino. 1997. Stability of aspartate aminotransferase from Sulfolobus solfataricus. Protein Eng. 10:237-248. - PubMed
-
- Bohm, G., and R. Jaenicke. 1994. Relevance of sequence statistics for the properties of extremophilic proteins. Int. J. Pept. Protein Res. 43:97-106. - PubMed
-
- Boyd, D. R., N. D. Sharma, and C. C. Allen. 2001. Aromatic dioxygenases: molecular biocatalysis and applications. Curr. Opin. Biotechnol. 12:564-573. - PubMed
-
- Carredano, E., A. Karlsson, B. Kauppi, D. Choudhury, R. E. Parales, J. V. Parales, K. Lee, D. T. Gibson, H. Eklund, and S. Ramaswamy. 2000. Substrate binding site of naphthalene 1,2-dioxygenase: functional implications of indole binding. J. Mol. Biol. 296:701-712. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
