

An expression-driven approach to the prediction of carbohydrate transport and utilization regulons in the hyperthermophilic bacterium Thermotoga maritima
- PMID: 16237010
- PMCID: PMC1272978
- DOI: 10.1128/JB.187.21.7267-7282.2005
An expression-driven approach to the prediction of carbohydrate transport and utilization regulons in the hyperthermophilic bacterium Thermotoga maritima
Abstract
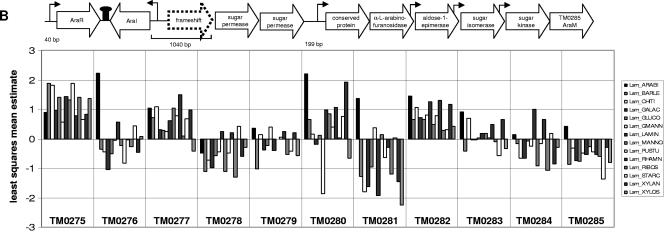
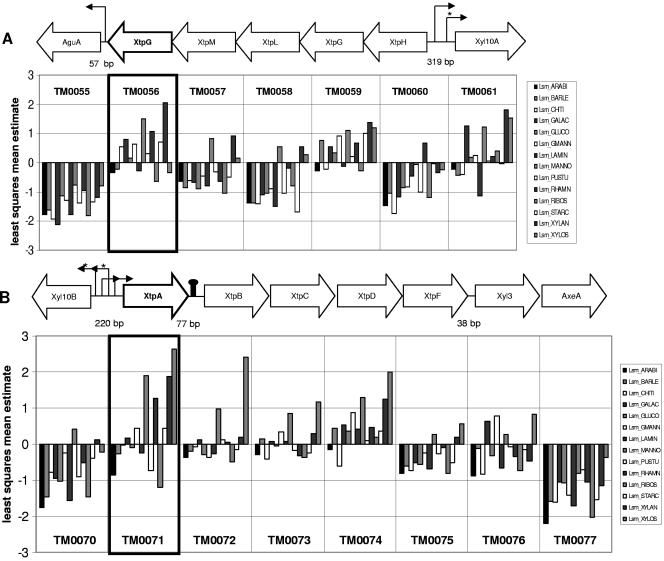
Comprehensive analysis of genome-wide expression patterns during growth of the hyperthermophilic bacterium Thermotoga maritima on 14 monosaccharide and polysaccharide substrates was undertaken with the goal of proposing carbohydrate specificities for transport systems and putative transcriptional regulators. Saccharide-induced regulons were predicted through the complementary use of comparative genomics, mixed-model analysis of genome-wide microarray expression data, and examination of upstream sequence patterns. The results indicate that T. maritima relies extensively on ABC transporters for carbohydrate uptake, many of which are likely controlled by local regulators responsive to either the transport substrate or a key metabolic degradation product. Roles in uptake of specific carbohydrates were suggested for members of the expanded Opp/Dpp family of ABC transporters. In this family, phylogenetic relationships among transport systems revealed patterns of possible duplication and divergence as a strategy for the evolution of new uptake capabilities. The presence of GC-rich hairpin sequences between substrate-binding proteins and other components of Opp/Dpp family transporters offers a possible explanation for differential regulation of transporter subunit genes. Numerous improvements to T. maritima genome annotations were proposed, including the identification of ABC transport systems originally annotated as oligopeptide transporters as candidate transporters for rhamnose, xylose, beta-xylan, and beta-glucans and identification of genes likely to encode proteins missing from current annotations of the pentose phosphate pathway. Beyond the information obtained for T. maritima, the present study illustrates how expression-based strategies can be used for improving genome annotation in other microorganisms, especially those for which genetic systems are unavailable.
Figures










References
-
- Bandlish, R. K., J. Michael Hess, K. L. Epting, C. Vieille, and R. M. Kelly. 2002. Glucose-to-fructose conversion at high temperatures with xylose (glucose) isomerases from Streptomyces murinus and two hyperthermophilic Thermotoga species. Biotechnol. Bioeng. 80:185-194. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
