

The homeodomain transcription factor Irx5 establishes the mouse cardiac ventricular repolarization gradient
- PMID: 16239150
- PMCID: PMC1480411
- DOI: 10.1016/j.cell.2005.08.004
The homeodomain transcription factor Irx5 establishes the mouse cardiac ventricular repolarization gradient
Abstract
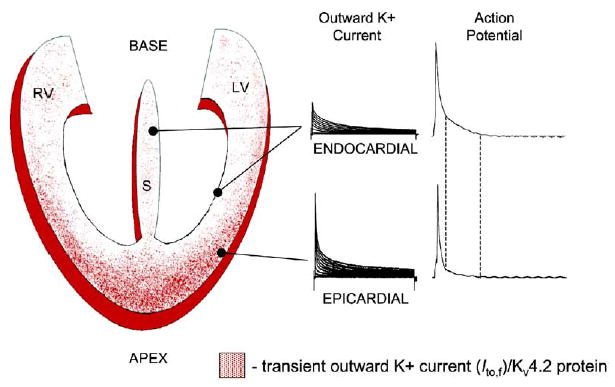
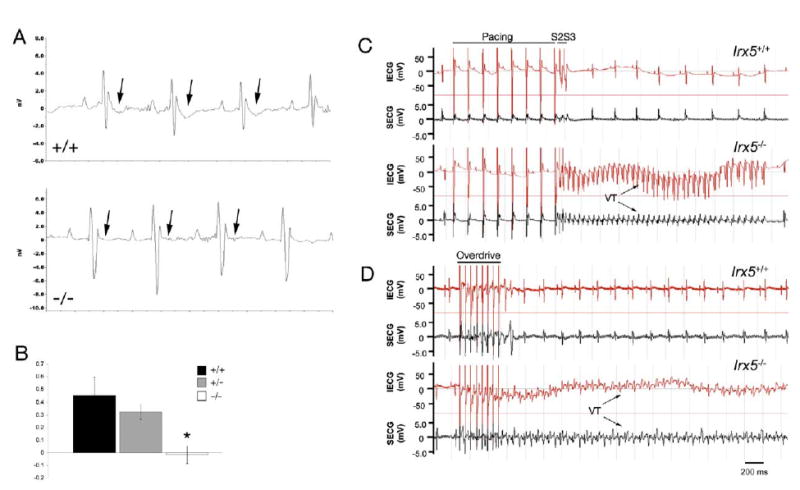
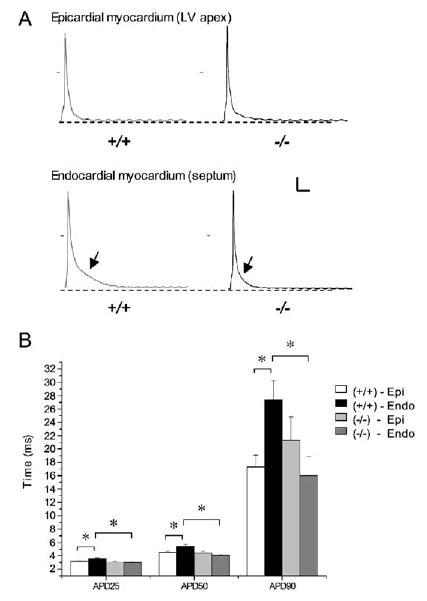
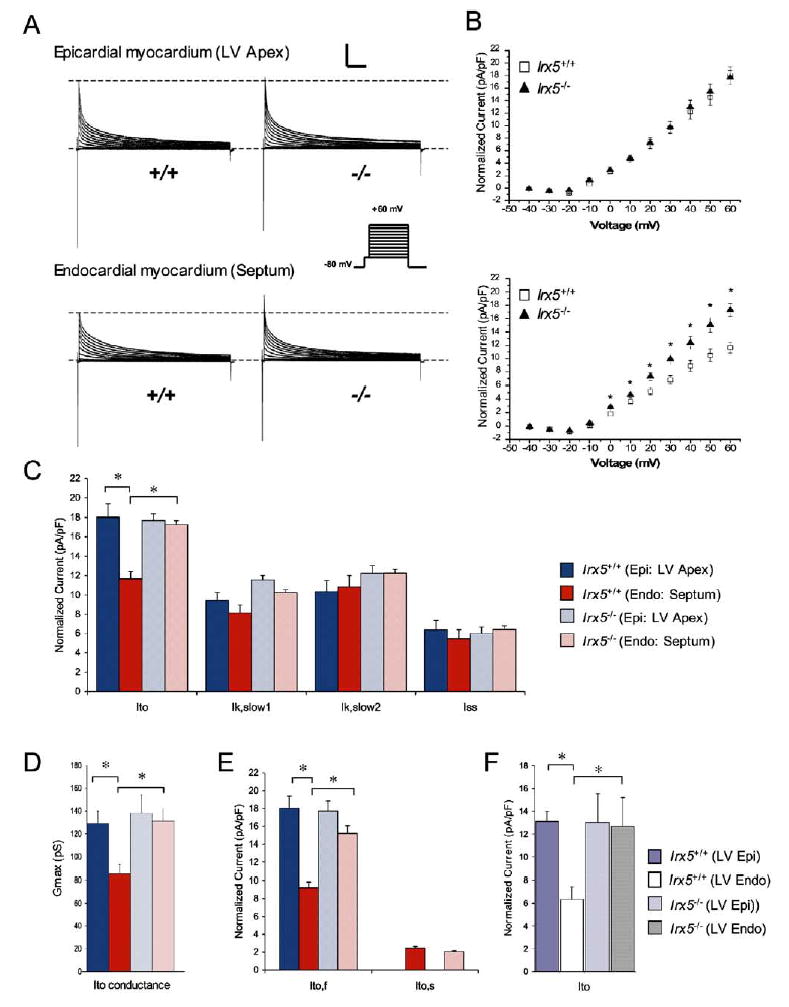
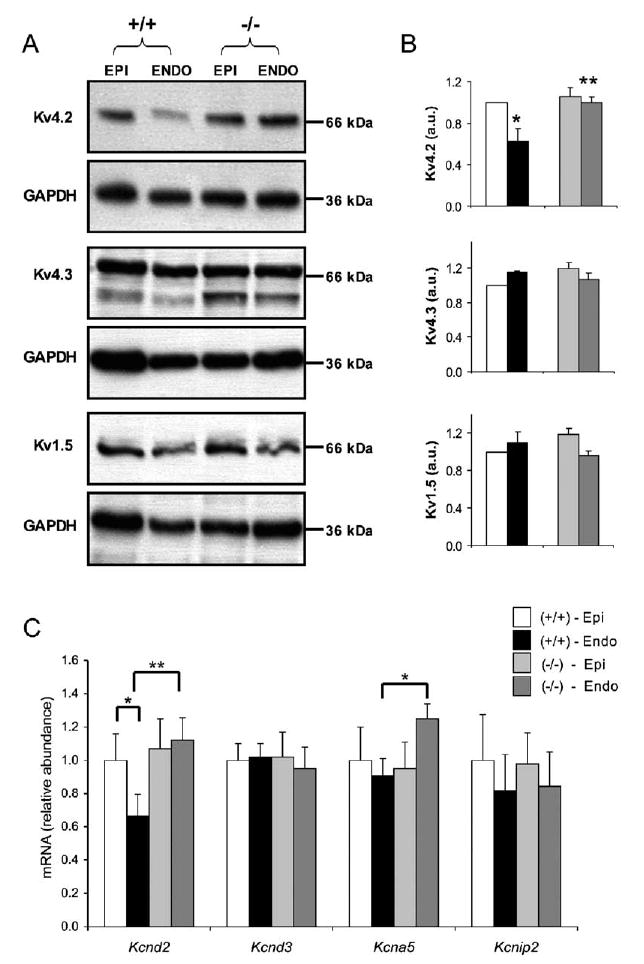
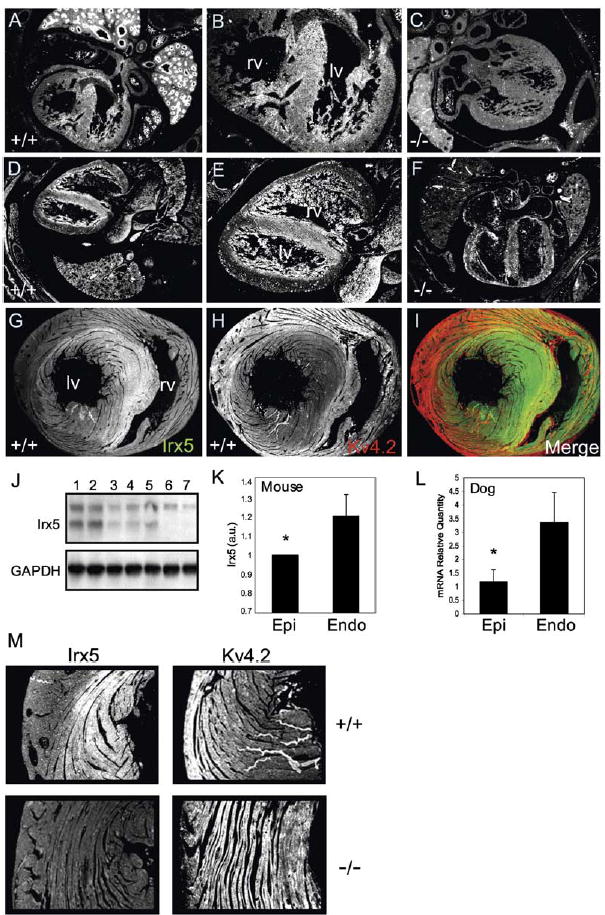
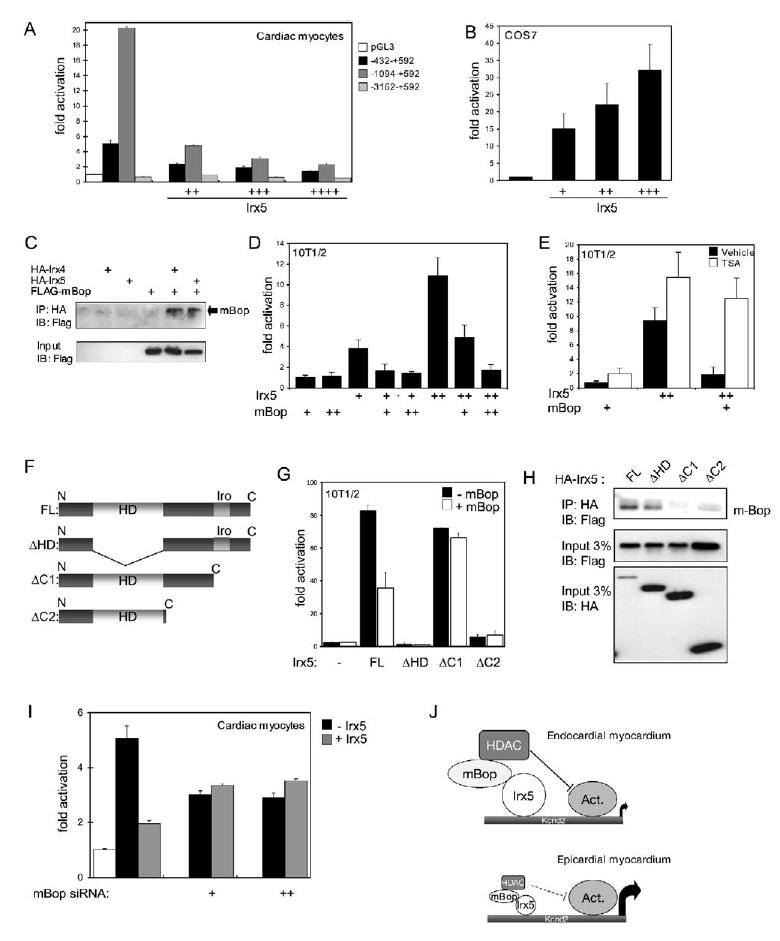
Rhythmic cardiac contractions depend on the organized propagation of depolarizing and repolarizing wavefronts. Repolarization is spatially heterogeneous and depends largely on gradients of potassium currents. Gradient disruption in heart disease may underlie susceptibility to fatal arrhythmias, but it is not known how this gradient is established. We show that, in mice lacking the homeodomain transcription factor Irx5, the cardiac repolarization gradient is abolished due to increased Kv4.2 potassium-channel expression in endocardial myocardium, resulting in a selective increase of the major cardiac repolarization current, I(to,f), and increased susceptibility to arrhythmias. Myocardial Irx5 is expressed in a gradient opposite that of Kv4.2, and Irx5 represses Kv4.2 expression by recruiting mBop, a cardiac transcriptional repressor. Thus, an Irx5 repressor gradient negatively regulates potassium-channel-gene expression in the heart, forming an inverse I(to,f) gradient that ensures coordinated cardiac repolarization while also preventing arrhythmias.
Figures
References
-
- Antzelevitch C. Cellular basis and mechanism underlying normal and abnormal myocardial repolarization and arrhythmogenesis. Ann Med. 2004;36 (Suppl 1):5–14. - PubMed
-
- Bao ZZ, Bruneau BG, Seidman JG, Seidman CE, Cepko CL. Irx4 regulates chamber-specific gene expression in the developing heart. Science. 1999;283:1161–1164. - PubMed
-
- Barry DM, Xu H, Schuessler RB, Nerbonne JM. Functional knockout of the transient outward current, long-QT syndrome, and cardiac remodeling in mice expressing a dominant-negative Kv4 alpha subunit. Circ Res. 1998;83:560–567. - PubMed
-
- Bruneau BG. Transcriptional regulation of vertebrate cardiac morphogenesis. Circ Res. 2002;90:509–519. - PubMed
-
- Bruneau BG, Bao ZZ, Tanaka M, Schott JJ, Izumo S, Cepko CL, Seidman JG, Seidman CE. Cardiac expression of the ventricle-specific homeobox gene Irx4 is modulated by Nkx2–5 and dHand. Dev Biol. 2000;217:266–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
