

Modulation by melatonin of glutamatergic synaptic transmission in the carp retina
- PMID: 16239269
- PMCID: PMC1464261
- DOI: 10.1113/jphysiol.2005.098798
Modulation by melatonin of glutamatergic synaptic transmission in the carp retina
Abstract
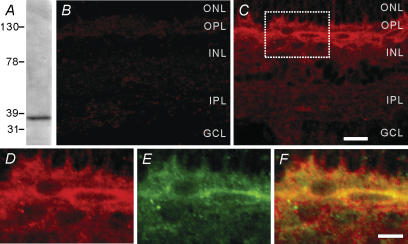
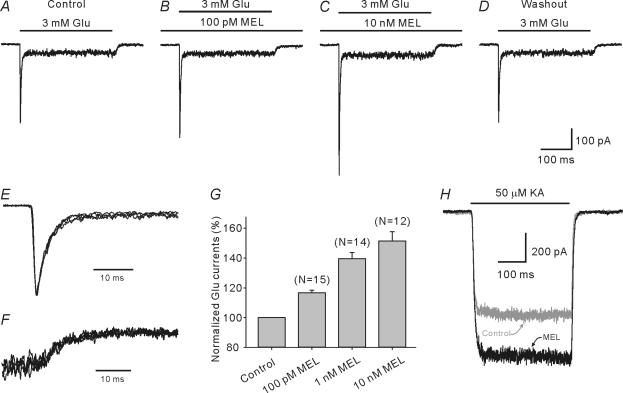
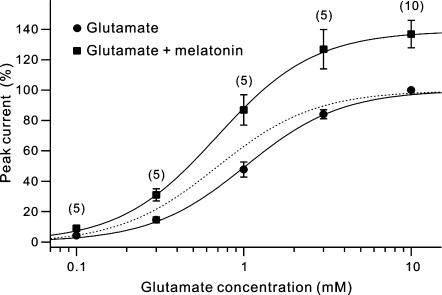
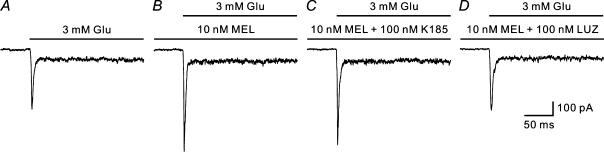
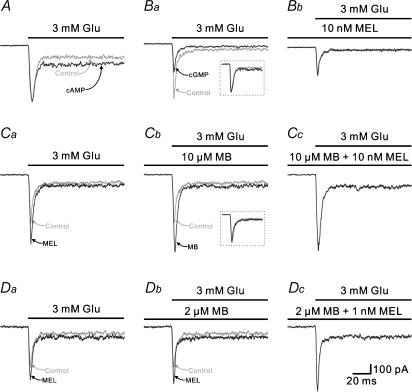
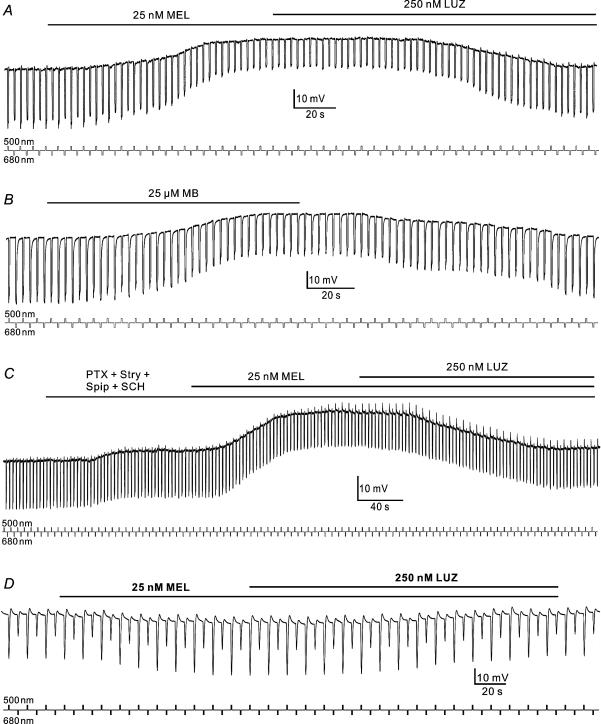
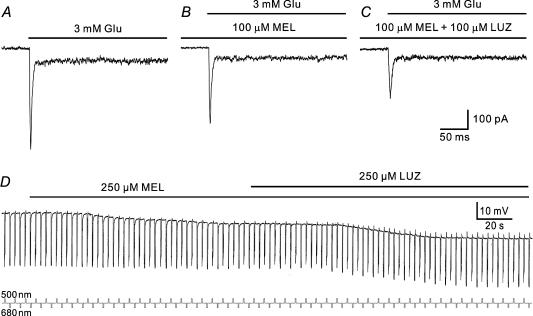
Melatonin is involved in a variety of physiological functions through activating specific receptors coupled to GTP-binding protein. Melatonin and its receptors are abundant in the retina. Here we show for the first time that melatonin modulates glutamatergic synaptic transmission from cones to horizontal cells (HCs) in carp retina. Immunocytochemical data revealed the expression of the MT1 receptor on carp HCs. Whole-cell recordings further showed that melatonin of physiological concentrations potentiated glutamate-induced currents from isolated cone-driven HCs (H1 cells) in a dose-dependent manner, by increasing the efficacy and apparent affinity of the glutamate receptor. The effects of melatonin were reversed by luzindole, but not by K 185, indicating the involvement of the MT1 receptor. Like melatonin, methylene blue (MB), a guanylate cyclase inhibitor, also potentiated the glutamate currents, but internal infusion of cGMP suppressed them. The effects of melatonin were not observed in cGMP-filled and MB-incubated HCs. These results suggest that the melatonin effects may be mediated by decreasing the intracellular concentration of cGMP. Consistent with these observations, melatonin depolarized the membrane potential of H1 cells and reduced their light responses, which could also be blocked by luzindole. These effects of melatonin persisted in the presence of the antagonists of receptors for dopamine, GABA and glycine, indicating a direct action of melatonin on H1 cells. Such modulation by melatonin of glutamatergic transmission from cones to HCs is thought to be in part responsible for circadian changes in light responsiveness of cone HCs in teleost retina.
Figures
Similar articles
-
Functional N-methyl-D-aspartate receptors are expressed in cone-driven horizontal cells in carp retina.Neurosignals. 2006-2007;15(4):174-9. doi: 10.1159/000096350. Epub 2006 Oct 17. Neurosignals. 2006. PMID: 17047395
-
Functional GABA(B) receptors are expressed at the cone photoreceptor terminals in bullfrog retina.Neuroscience. 2005;132(1):103-13. doi: 10.1016/j.neuroscience.2004.12.024. Neuroscience. 2005. PMID: 15780470
-
Expression of connexin 35/36 in retinal horizontal and bipolar cells of carp.Neuroscience. 2009 Dec 15;164(3):1161-9. doi: 10.1016/j.neuroscience.2009.09.035. Epub 2009 Sep 22. Neuroscience. 2009. PMID: 19778581
-
The involvement of glutamate-gated channels in negative feedback from horizontal cells to cones.Prog Brain Res. 2005;147:219-29. doi: 10.1016/S0079-6123(04)47017-4. Prog Brain Res. 2005. PMID: 15581709 Review.
-
Neuromodulatory role of melatonin in retinal information processing.Prog Retin Eye Res. 2013 Jan;32:64-87. doi: 10.1016/j.preteyeres.2012.07.003. Epub 2012 Aug 16. Prog Retin Eye Res. 2013. PMID: 22986412 Review.
Cited by
-
Environmental Cycles, Melatonin, and Circadian Control of Stress Response in Fish.Front Endocrinol (Lausanne). 2019 Jun 11;10:279. doi: 10.3389/fendo.2019.00279. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31244768 Free PMC article. Review.
-
Gut melatonin response to microbial infection in carp Catla catla.Fish Physiol Biochem. 2016 Apr;42(2):579-92. doi: 10.1007/s10695-015-0161-7. Epub 2015 Nov 13. Fish Physiol Biochem. 2016. PMID: 26563281
-
Melatonin potentiates glycine currents through a PLC/PKC signalling pathway in rat retinal ganglion cells.J Physiol. 2010 Jul 15;588(Pt 14):2605-19. doi: 10.1113/jphysiol.2010.187641. Epub 2010 Jun 2. J Physiol. 2010. PMID: 20519319 Free PMC article.
-
Cone Viability Is Affected by Disruption of Melatonin Receptors Signaling.Invest Ophthalmol Vis Sci. 2016 Jan 1;57(1):94-104. doi: 10.1167/iovs.15-18235. Invest Ophthalmol Vis Sci. 2016. PMID: 26780313 Free PMC article.
-
Melatonin potentiates rod signals to ON type bipolar cells in fish retina.J Physiol. 2008 Jun 1;586(11):2683-94. doi: 10.1113/jphysiol.2008.152959. Epub 2008 Apr 3. J Physiol. 2008. PMID: 18388138 Free PMC article.
References
-
- Alonso-Gomez AL, Valenciano AI, Alonso-Bedate M, Delgado MJ. Melatonin synthesis in the greenfrog retina in culture. I. Modulation by the light/dark cycle, forskolin and inhibitors of protein synthesis. Life Sci. 2000;66:675–685. - PubMed
-
- Besharse JC, Iuvone PM. Circadian clock in Xenopus eye controlling retinal serotonin N-acetyltransferase. Nature. 1983;305:133–135. - PubMed
-
- Brzezinski A. Melatonin in humans. N Engl J Med. 1997;336:186–195. - PubMed
-
- Cahill GM. Circadian regulation of melatonin production in cultured zebrafish pineal and retina. Brain Res. 1996;708:177–181. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous