

Pathogenesis of Yersinia pestis infection in BALB/c mice: effects on host macrophages and neutrophils
- PMID: 16239508
- PMCID: PMC1273833
- DOI: 10.1128/IAI.73.11.7142-7150.2005
Pathogenesis of Yersinia pestis infection in BALB/c mice: effects on host macrophages and neutrophils
Abstract
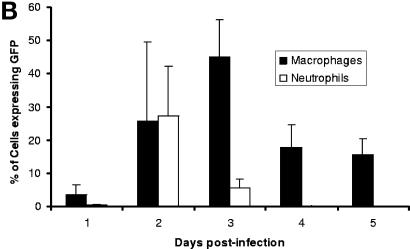
The pathogenesis of infection with Yersinia pestis, the causative agent of plague, was examined following subcutaneous infection of BALB/c mice with a fully virulent strain expressing green fluorescent protein. Plate culturing, flow cytometry, and laser confocal microscopy of spleen homogenates throughout infection revealed three discernible stages of infection. The early phase was characterized by the presence of a small number of intracellular bacteria mostly within CD11b+ macrophages and Ly-6G+ neutrophils. These bacteria were not viable, as determined by plate culturing of spleen homogenates, until day 2 postinfection. Between days 2 and 4 postinfection, a plateau phase was observed, with bacterial burdens of 10(3) to 10(4) CFU per spleen. Flow cytometric analysis revealed that there was even distribution of Y. pestis within both CD11b+ macrophage and Ly-6G+ neutrophil populations on day 2 postinfection. However, from day 3 postinfection onward, intracellular bacteria were observed exclusively within splenic CD11b+ macrophages. The late phase of infection, between days 4 and 5 postinfection, was characterized by a rapid increase in bacterial numbers, as well as escape of bacteria into the extracellular compartment. Annexin V staining of spleens indicated that a large proportion of splenic neutrophils underwent rapid apoptosis on days 1 and 2 postinfection. Fewer macrophages underwent apoptosis during the same period. Our data suggest that during the early stages of Y. pestis infection, splenic neutrophils are responsible for limiting the growth of Y. pestis and that splenic macrophages provide safe intracellular shelters within which Y. pestis is able to grow and escape during the later stages of infection. This macrophage compliance can be overcome in vitro by stimulation with a combination of gamma interferon and tumor necrosis factor alpha.
Figures





References
-
- Andrew, P. W., J. P. S. Jackett, and D. B. Lowrie. 1985. Killing and degradation of microorganisms by macrophages, p. 311-335. In R. T. Dean, W. Jessop, and J. W. Dean (ed.), Mononuclear phagocytes: physiology and pathology. Elsevier Biomedical Press, Amsterdam, The Netherlands.
-
- Cavanaugh, D., and R. Randall. 1959. The role of multiplication of Pasteurella pestis in mononuclear phagocytes in the pathogenesis of flea-borne plague. J. Immunol. 83:348-363. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
