

Toxoplasma gondii acyl-lipid metabolism: de novo synthesis from apicoplast-generated fatty acids versus scavenging of host cell precursors
- PMID: 16246004
- PMCID: PMC1386017
- DOI: 10.1042/BJ20050609
Toxoplasma gondii acyl-lipid metabolism: de novo synthesis from apicoplast-generated fatty acids versus scavenging of host cell precursors
Abstract
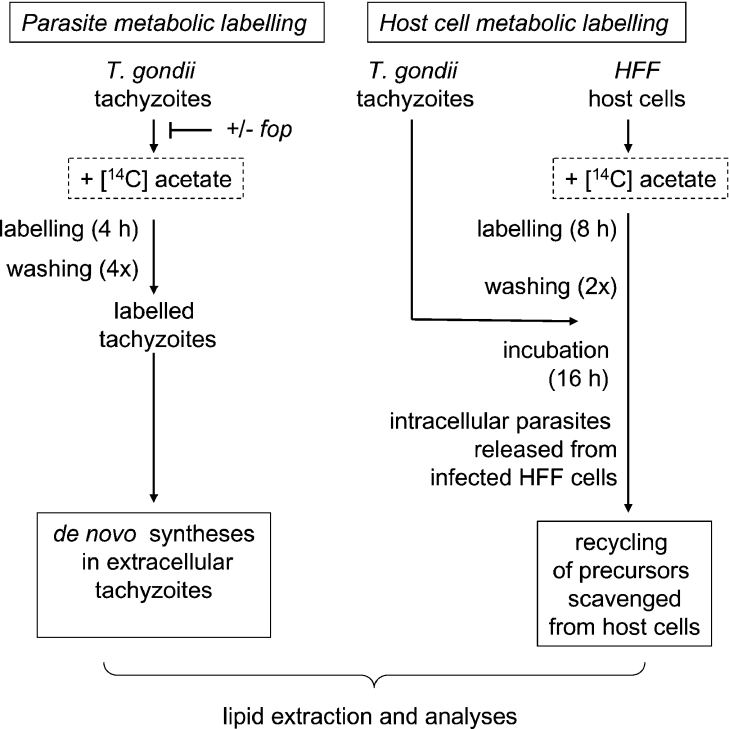
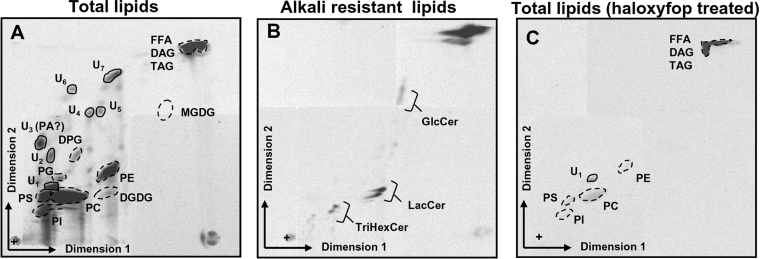
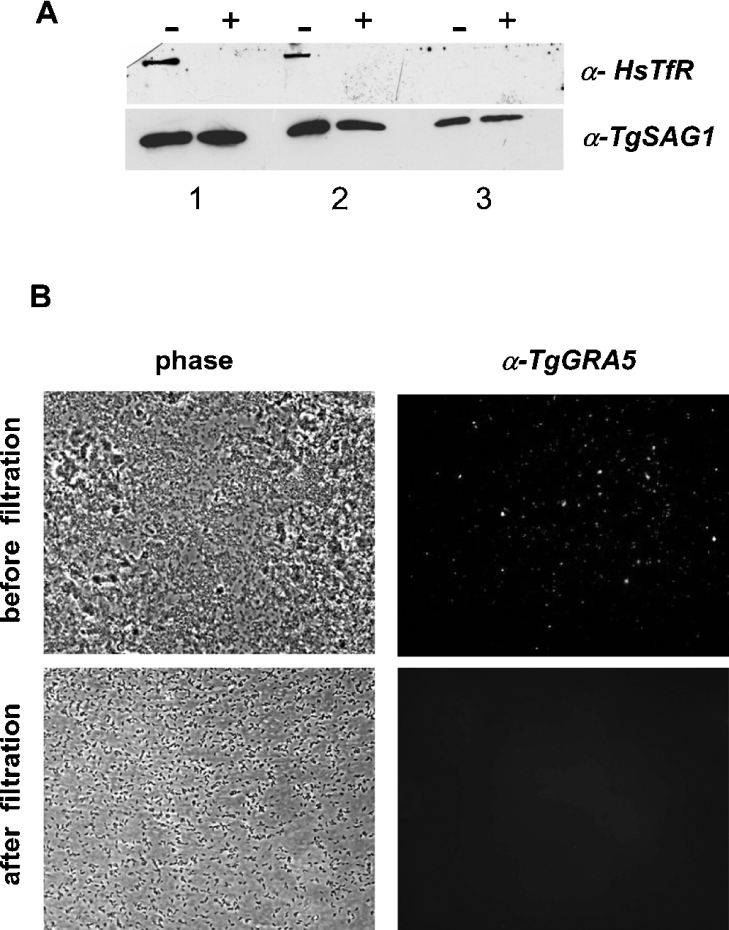
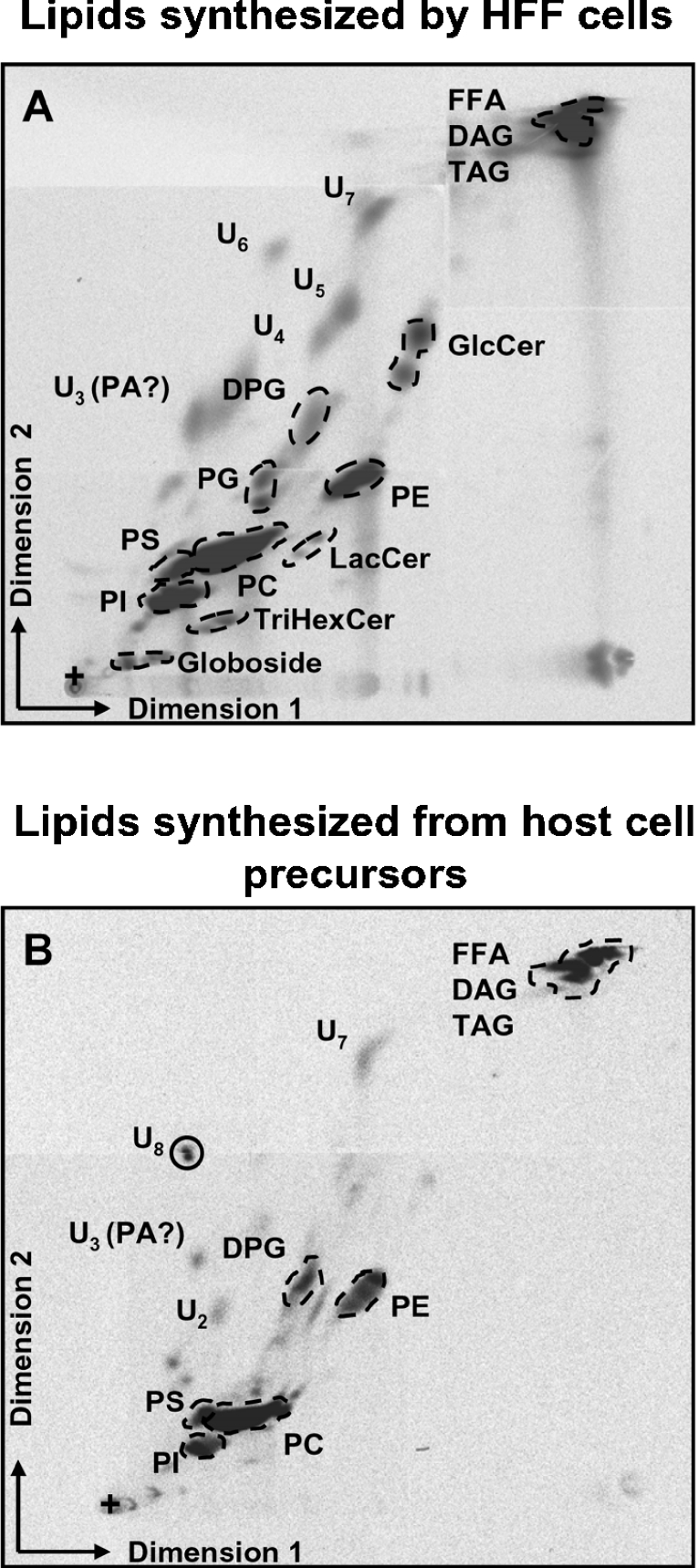
Toxoplasma gondii is an obligate intracellular parasite that contains a relic plastid, called the apicoplast, deriving from a secondary endosymbiosis with an ancestral alga. Metabolic labelling experiments using [14C]acetate led to a substantial production of numerous glycero- and sphingo-lipid classes in extracellular tachyzoites. Syntheses of all these lipids were affected by the herbicide haloxyfop, demonstrating that their de novo syntheses necessarily required a functional apicoplast fatty acid synthase II. The complex metabolic profiles obtained and a census of glycerolipid metabolism gene candidates indicate that synthesis is probably scattered in the apicoplast membranes [possibly for PA (phosphatidic acid), DGDG (digalactosyldiacylglycerol) and PG (phosphatidylglycerol)], the endoplasmic reticulum (for major phospholipid classes and ceramides) and mitochondria (for PA, PG and cardiolipid). Based on a bioinformatic analysis, it is proposed that apicoplast produced acyl-ACP (where ACP is acyl-carrier protein) is transferred to glycerol-3-phosphate for apicoplast glycerolipid synthesis. Acyl-ACP is also probably transported outside the apicoplast stroma and irreversibly converted into acyl-CoA. In the endoplasmic reticulum, acyl-CoA may not be transferred to a three-carbon backbone by an enzyme similar to the cytosolic plant glycerol-3-phosphate acyltransferase, but rather by a dual glycerol-3-phosphate/dihydroxyacetone-3-phosphate acyltransferase like in animal and yeast cells. We further showed that intracellular parasites could also synthesize most of their lipids from scavenged host cell precursors. The observed appearance of glycerolipids specific to either the de novo pathway in extracellular parasites (unknown glycerolipid 1 and the plant like DGDG), or the intracellular stages (unknown glycerolipid 8), may explain the necessary coexistence of both de novo parasitic acyl-lipid synthesis and recycling of host cell compounds.
Figures
Similar articles
-
Apicoplast fatty acid synthesis is essential for organelle biogenesis and parasite survival in Toxoplasma gondii.Proc Natl Acad Sci U S A. 2006 Aug 29;103(35):13192-7. doi: 10.1073/pnas.0603391103. Epub 2006 Aug 18. Proc Natl Acad Sci U S A. 2006. PMID: 16920791 Free PMC article.
-
Comprehensive Characterization of Toxoplasma Acyl Coenzyme A-Binding Protein TgACBP2 and Its Critical Role in Parasite Cardiolipin Metabolism.mBio. 2018 Oct 23;9(5):e01597-18. doi: 10.1128/mBio.01597-18. mBio. 2018. PMID: 30352931 Free PMC article.
-
The acyl-CoA synthetase TgACS1 allows neutral lipid metabolism and extracellular motility in Toxoplasma gondii through relocation via its peroxisomal targeting sequence (PTS) under low nutrient conditions.mBio. 2024 Apr 10;15(4):e0042724. doi: 10.1128/mbio.00427-24. Epub 2024 Mar 19. mBio. 2024. PMID: 38501871 Free PMC article.
-
Lipid synthesis in protozoan parasites: a comparison between kinetoplastids and apicomplexans.Prog Lipid Res. 2013 Oct;52(4):488-512. doi: 10.1016/j.plipres.2013.06.003. Epub 2013 Jul 1. Prog Lipid Res. 2013. PMID: 23827884 Free PMC article. Review.
-
Phosphatidic acid synthesis in bacteria.Biochim Biophys Acta. 2013 Mar;1831(3):495-502. doi: 10.1016/j.bbalip.2012.08.018. Epub 2012 Aug 30. Biochim Biophys Acta. 2013. PMID: 22981714 Free PMC article. Review.
Cited by
-
Omega-3 Polyunsaturated Fatty Acids Prevent Toxoplasma gondii Infection by Inducing Autophagy via AMPK Activation.Nutrients. 2019 Sep 6;11(9):2137. doi: 10.3390/nu11092137. Nutrients. 2019. PMID: 31500218 Free PMC article.
-
Further aspects of Toxoplasma gondii elimination in the presence of metals.Parasitol Res. 2018 Apr;117(4):1245-1256. doi: 10.1007/s00436-018-5806-x. Epub 2018 Feb 18. Parasitol Res. 2018. PMID: 29455419
-
Transcriptomic analyses reveal distinct response of porcine macrophages to Toxoplasma gondii infection.Parasitol Res. 2020 Jun;119(6):1819-1828. doi: 10.1007/s00436-020-06677-5. Epub 2020 May 13. Parasitol Res. 2020. PMID: 32399721
-
Plasmodium DEH is ER-localized and crucial for oocyst mitotic division during malaria transmission.Life Sci Alliance. 2020 Oct 26;3(12):e202000879. doi: 10.26508/lsa.202000879. Print 2020 Dec. Life Sci Alliance. 2020. PMID: 33106323 Free PMC article.
-
The influence of exposure to Toxoplasma Gondii on host lipid metabolism.BMC Infect Dis. 2020 Jun 15;20(1):415. doi: 10.1186/s12879-020-05138-9. BMC Infect Dis. 2020. PMID: 32539811 Free PMC article.
References
-
- Dubey J. P. Toxoplasma, Hammondia, Besnoitia, Sarcocystis, and other tissue cyst-forming coccidian of man and animals. In: Kreier J. P., editor. Parasitic Protozoa, vol. III. New York: Academic Press; 1977. pp. 101–237.
-
- Luft B. J., Hafner R., Korzun A. H., Leport C., Antoniskis D., Bosler E. M., Bourland D. D., Uttamchandani R., Fuhrer J., Jacobson J., et al. Toxoplasmic encephalitis in patients with the acquired immunodeficiency syndrome. N. Engl. J. Med. 1993;329:995–1000. - PubMed
-
- Wong S. Y., Remington J. S. Toxoplasmosis in pregnancy. Clin. Infect. Dis. 1994;18:853–862. - PubMed
-
- Wilson R. J. M., Williamson D. H., Preiser P. Malaria and other apicomplexans: the ‘plant’ connection. Infect. Agents Dis. 1994;3:29–37. - PubMed
-
- Wilson R. J. M., Denny P. W., Preiser P. R., Rangachari K., Roberts K., Roy A., Whyte A., Strath M., Moore D. J., Moore P. W., et al. Complete gene map of the plastid-like DNA of the malaria parasite Plasmodium falciparum. J. Mol. Biol. 1996;261:155–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources