

The response of autologous T cells to a human melanoma is dominated by mutated neoantigens
- PMID: 16247014
- PMCID: PMC1266037
- DOI: 10.1073/pnas.0500090102
The response of autologous T cells to a human melanoma is dominated by mutated neoantigens
Abstract
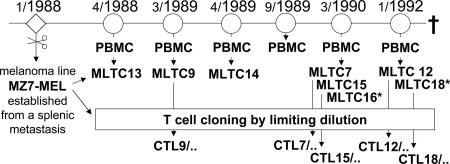
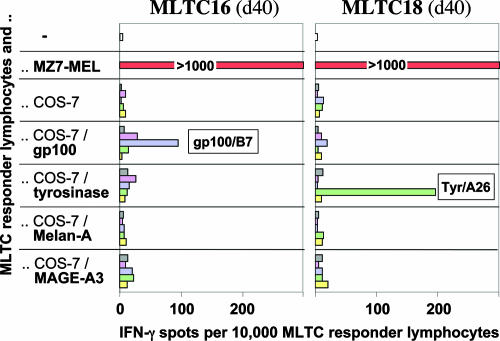
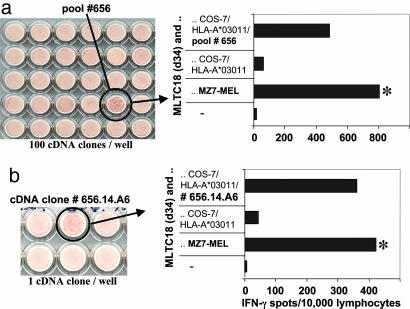
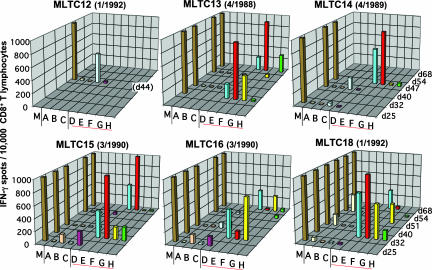
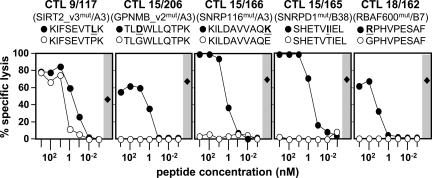
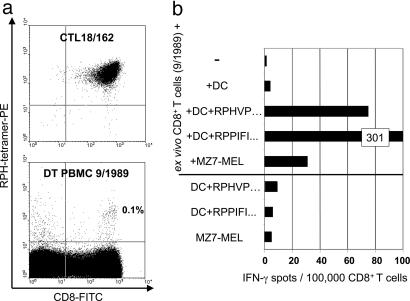
Our understanding of pathways leading to antitumor immunity may depend on an undistorted knowledge of the primary antigenic targets of patients' autologous T cell responses. In the melanoma model derived from patient DT, we applied cryopreserved short-term autologous mixed lymphocyte-tumor cell cultures (MLTCs) in combination with an IFN-gamma enzyme-linked immunospot (ELISPOT) assay to cDNA expression screening. We identified three previously unknown peptides processed from melanosomal proteins tyrosinase (presented by HLA-A(*)2601 and -B(*)3801) and gp100 (presented by HLA-B(*)07021) and five neoantigens generated by somatic point mutations in the patient's melanoma. The mutations were found in the genes SIRT2, GPNMB, SNRP116, SNRPD1, and RBAF600. Peptides containing the mutated residues were presented by HLA-A(*)03011, -B(*)07021, and -B(*)3801. Mutation-induced functional impairment was so far demonstrated for SIRT2. Within MLTC responder populations that were independently expanded from the patient's peripheral blood lymphocytes of different years, T cells against mutated epitopes clearly predominated. These results document a high degree of individuality for the cellular antitumor response and support the need for individualizing the monitoring and therapeutic approaches to the primary targets of the autologous T cell response, which may finally lead to a more effective cancer immunotherapy.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
