

The mechanisms and dynamics of (alpha)v(beta)3 integrin clustering in living cells
- PMID: 16247034
- PMCID: PMC2171205
- DOI: 10.1083/jcb.200503017
The mechanisms and dynamics of (alpha)v(beta)3 integrin clustering in living cells
Abstract
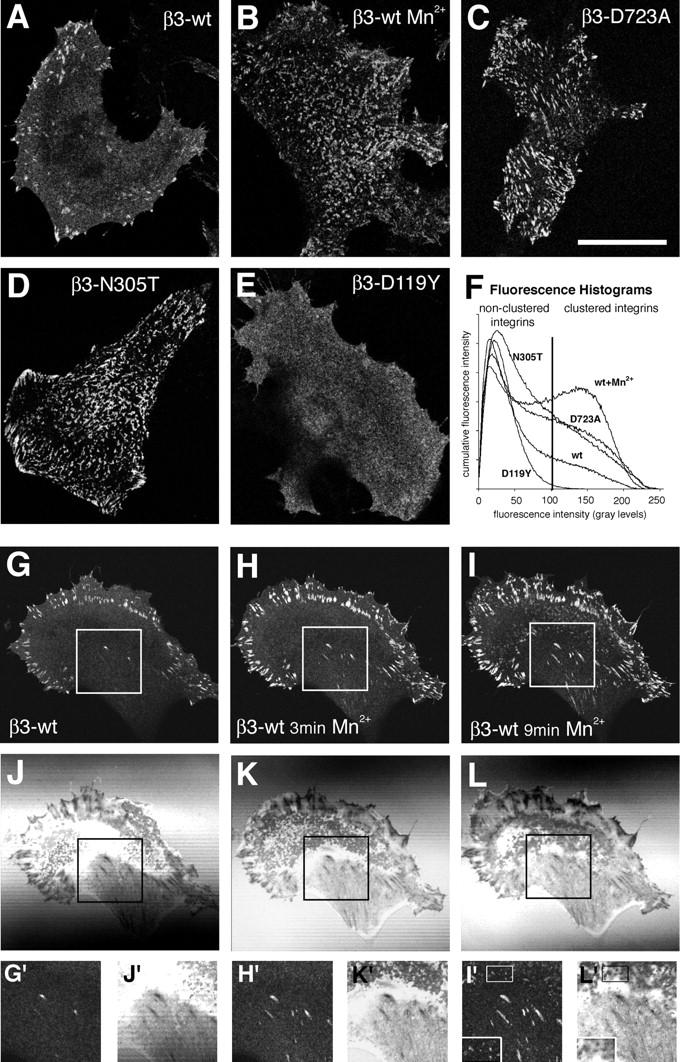
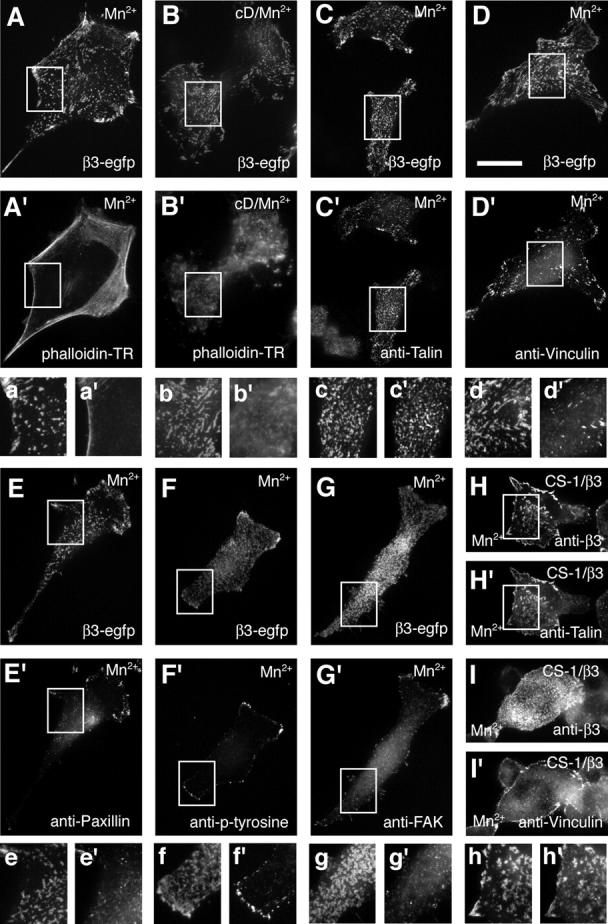
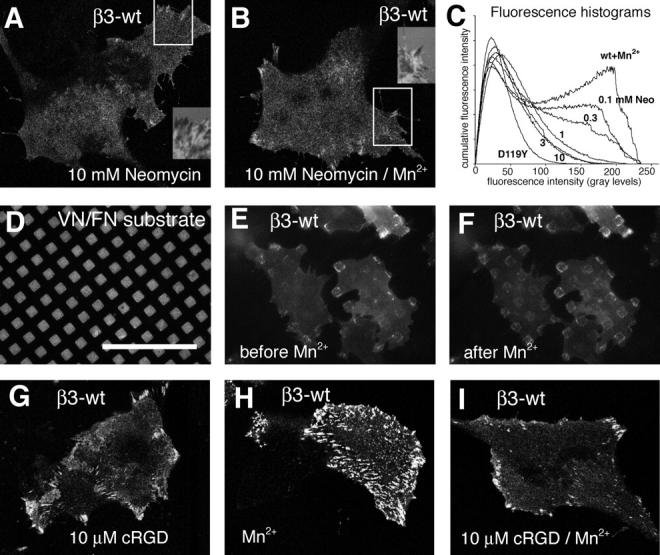
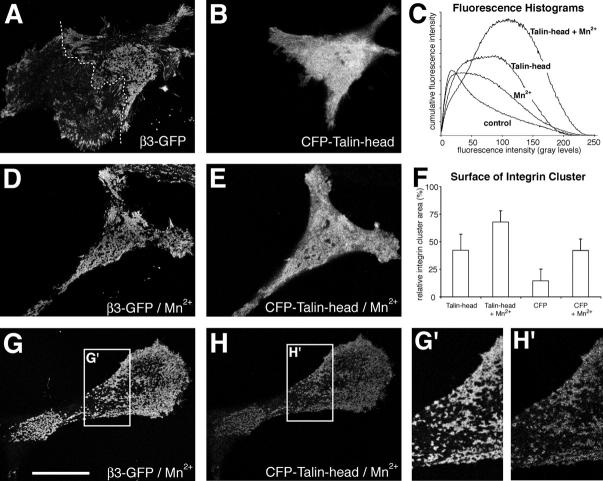
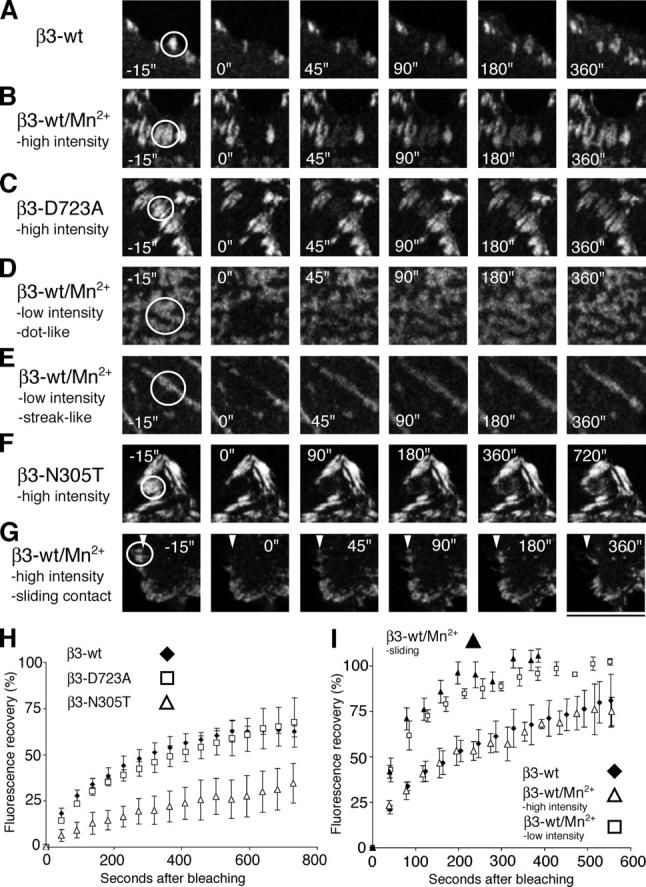
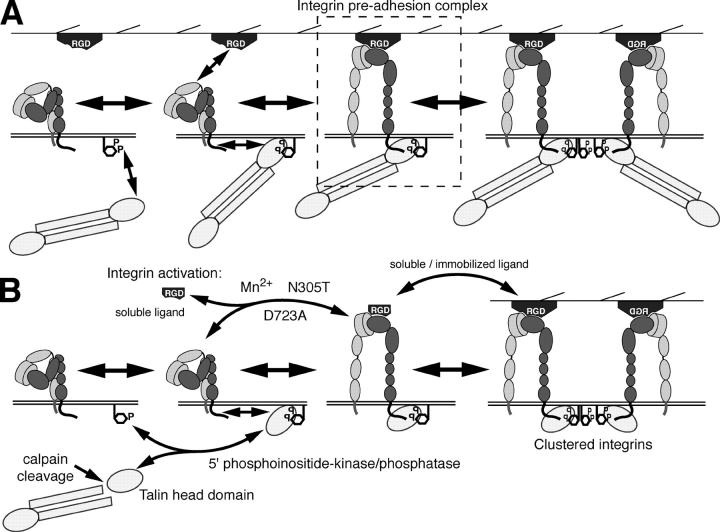
During cell migration, the physical link between the extracellular substrate and the actin cytoskeleton mediated by receptors of the integrin family is constantly modified. We analyzed the mechanisms that regulate the clustering and incorporation of activated alphavbeta3 integrins into focal adhesions. Manganese (Mn2+) or mutational activation of integrins induced the formation of de novo F-actin-independent integrin clusters. These clusters recruited talin, but not other focal adhesion adapters, and overexpression of the integrin-binding head domain of talin increased clustering. Integrin clustering required immobilized ligand and was prevented by the sequestration of phosphoinositole-4,5-bisphosphate (PI(4,5)P2). Fluorescence recovery after photobleaching analysis of Mn(2+)-induced integrin clusters revealed increased integrin turnover compared with mature focal contacts, whereas stabilization of the open conformation of the integrin ectodomain by mutagenesis reduced integrin turnover in focal contacts. Thus, integrin clustering requires the formation of the ternary complex consisting of activated integrins, immobilized ligands, talin, and PI(4,5)P2. The dynamic remodeling of this ternary complex controls cell motility.
Figures
References
-
- Arbuzova, A., K. Martushova, G. Hangyas-Mihalyne, A.J. Morris, S. Ozaki, G.D. Prestwich, and S. McLaughlin. 2000. Fluorescently labeled neomycin as a probe of phosphatidylinositol-4,5-bisphosphate in membranes. Biochim. Biophys. Acta. 1464:35–48. - PubMed
-
- Barsukov, I.L., A. Prescot, N. Bate, B. Patel, D.N. Floyd, N. Bhanji, C.R. Bagshaw, K. Letinic, G. Di Paolo, P. De Camilli, et al. 2003. Phosphatidylinositol phosphate kinase type 1gamma and beta1-integrin cytoplasmic domain bind to the same region in the talin FERM domain. J. Biol. Chem. 278:31202–31209. - PubMed
-
- Calderwood, D.A., B. Yan, J.M. de Pereda, B.G. Alvarez, Y. Fujioka, R.C. Liddington, and M.H. Ginsberg. 2002. The phosphotyrosine binding-like domain of talin activates integrins. J. Biol. Chem. 277:21749–21758. - PubMed
-
- Chen, J., A. Salas, and T.A. Springer. 2003. Bistable regulation of integrin adhesiveness by a bipolar metal ion cluster. Nat. Struct. Biol. 10:995–1001. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
