

Unique suppression of prostaglandin H synthase-2 expression by inhibition of histone deacetylation, specifically in human amnion but not adjacent choriodecidua
- PMID: 16251350
- PMCID: PMC1345689
- DOI: 10.1091/mbc.e05-08-0818
Unique suppression of prostaglandin H synthase-2 expression by inhibition of histone deacetylation, specifically in human amnion but not adjacent choriodecidua
Abstract
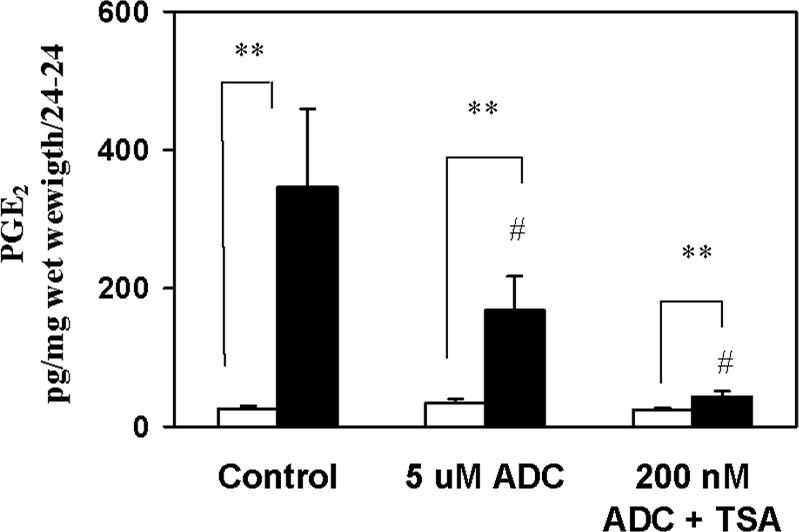
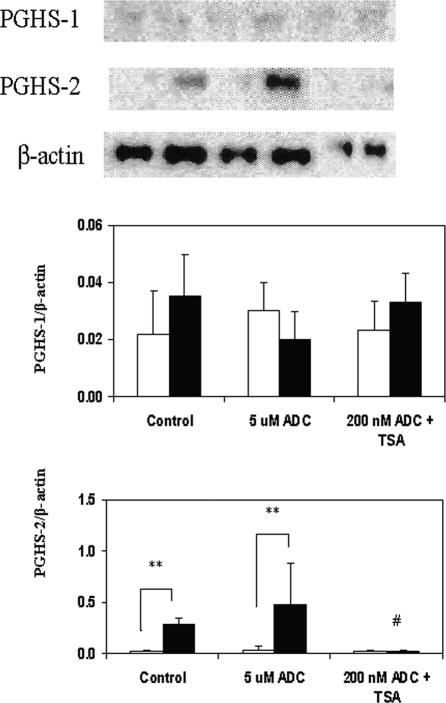
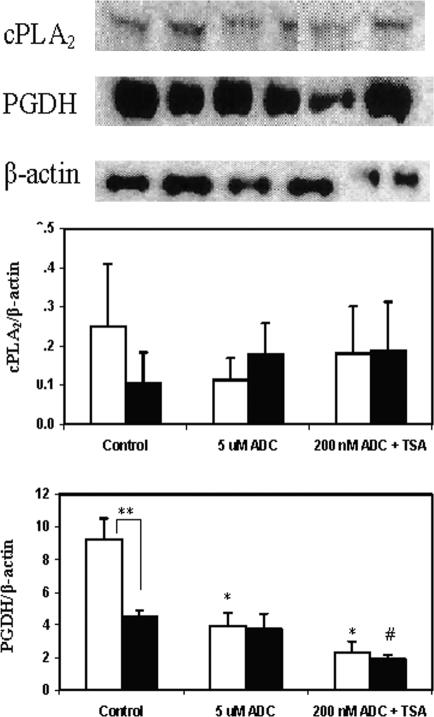
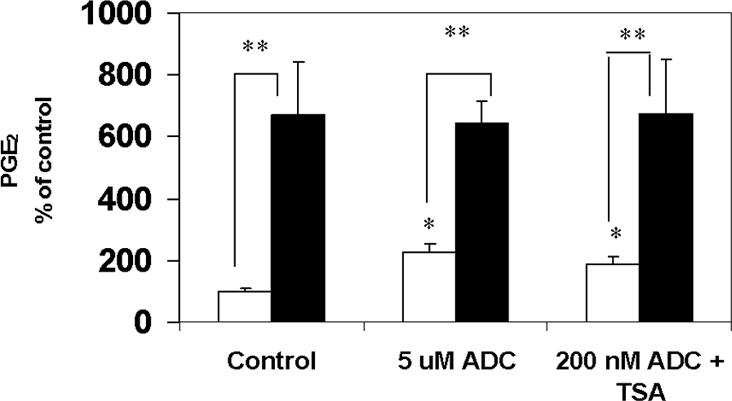
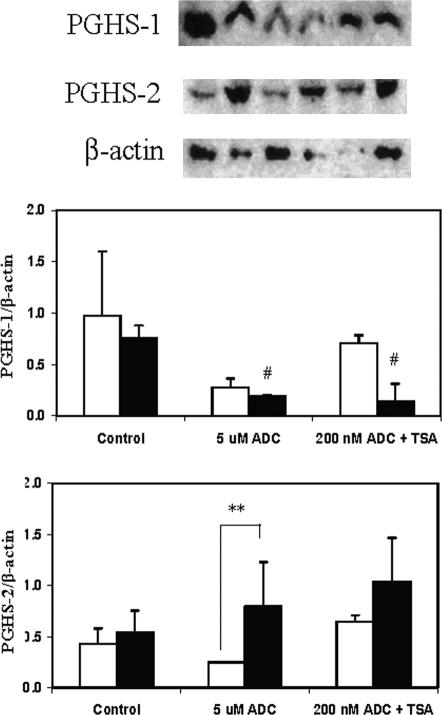
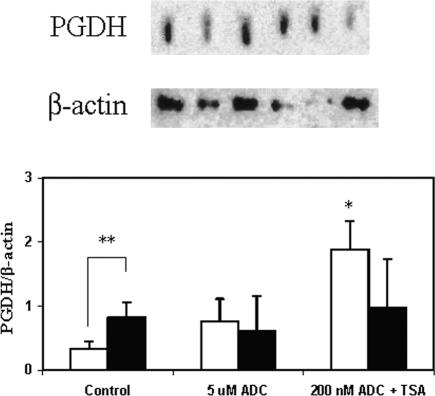
The key molecular regulatory mechanisms that govern and coordinate the molecular alterations that underpin the process of human labor remain incompletely understood although enhanced intrauterine prostaglandin production is known to be requisite. Studies from cancer tissues have indicated that at least one key enzyme of prostaglandin biosynthesis can have its activity severely reduced by increased histone deacetylation and enhanced DNA methylation status. We have advanced the hypothesis that similar regulation may occur in intrauterine tissues during pregnancy to prevent inadvertent activation of this powerful initiating signal by dampening responses to premature activation by agents such as cytokines. Our studies have shown that responsiveness of amnion, a key intrauterine tissue, to interleukin-1beta is abrogated by inhibition of histone deacetylation, whereas PGDH amounts were increased basally. The findings do integrate well with others concerning progesterone (inhibitory) actions such that a decrease in the level of histone acetylation in human gestational tissues near term might herald a coordinated series of events that all result in a positive drive for parturition. Hence, a new level of regulatory action and potential therapeutic targets for pathologies such as preterm labor can flow from these findings.
Figures
Similar articles
-
Prostaglandin H synthase-2 gene regulation in the amnion at labour: histone acetylation and nuclear factor kappa B binding to the promoter in vivo.Mol Hum Reprod. 2008 Jan;14(1):53-9. doi: 10.1093/molehr/gam086. Epub 2008 Jan 18. Mol Hum Reprod. 2008. PMID: 18204072
-
PTGS2 (prostaglandin endoperoxide synthase-2) expression in term human amnion in vivo involves rapid mRNA turnover, polymerase-II 5'-pausing, and glucocorticoid transrepression.Endocrinology. 2011 May;152(5):2113-22. doi: 10.1210/en.2010-1327. Epub 2011 Mar 8. Endocrinology. 2011. PMID: 21385935
-
Cannabinoids stimulate prostaglandin production by human gestational tissues through a tissue- and CB1-receptor-specific mechanism.Am J Physiol Endocrinol Metab. 2008 Feb;294(2):E352-6. doi: 10.1152/ajpendo.00495.2007. Epub 2007 Nov 27. Am J Physiol Endocrinol Metab. 2008. PMID: 18042663
-
Inhibition of lysyl oxidase by prostaglandin E2 via EP2/EP4 receptors in human amnion fibroblasts: Implications for parturition.Mol Cell Endocrinol. 2016 Mar 15;424:118-27. doi: 10.1016/j.mce.2016.01.022. Epub 2016 Jan 27. Mol Cell Endocrinol. 2016. PMID: 26826430
-
Prostaglandin dehydrogenase and the initiation of labor.J Perinat Med. 1999;27(1):26-34. doi: 10.1515/JPM.1999.003. J Perinat Med. 1999. PMID: 10343931 Review.
Cited by
-
Effectiveness of a federal healthy start program in reducing the impact of particulate air pollutants on feto-infant morbidity outcomes.Matern Child Health J. 2012 Nov;16(8):1602-11. doi: 10.1007/s10995-011-0854-1. Matern Child Health J. 2012. PMID: 21769586
-
Regulation of MT1-MMP/MMP-2/TIMP-2 axis in human placenta.J Inflamm Res. 2015 Oct 3;8:193-200. doi: 10.2147/JIR.S88039. eCollection 2015. J Inflamm Res. 2015. PMID: 26491367 Free PMC article.
-
The use of high-dimensional biology (genomics, transcriptomics, proteomics, and metabolomics) to understand the preterm parturition syndrome.BJOG. 2006 Dec;113 Suppl 3(Suppl 3):118-35. doi: 10.1111/j.1471-0528.2006.01150.x. BJOG. 2006. PMID: 17206980 Free PMC article. Review.
-
Epigenetic regulation of cytokine production in human amnion and villous placenta.Mediators Inflamm. 2012;2012:159709. doi: 10.1155/2012/159709. Epub 2012 May 14. Mediators Inflamm. 2012. PMID: 22665947 Free PMC article.
-
Regulation of TIMP-1 in Human Placenta and Fetal Membranes by lipopolysaccharide and demethylating agent 5-aza-2'-deoxycytidine.Reprod Biol Endocrinol. 2015 Dec 21;13:136. doi: 10.1186/s12958-015-0132-y. Reprod Biol Endocrinol. 2015. PMID: 26691525 Free PMC article.
References
-
- Akhtar, M., Cheng, Y., Magno, R. M., Ashktorab, H., Smoot, D. T., Meltzer, S. J., and Wilson, K. T. (2001). Promoter methylation regulates helicobacter pylori-stimulated cyclooxygenase-2 expression in gastric epithelial cells. Cancer Res. 61, 2399-2403. - PubMed
-
- Blumenstein, M., Bowen-Shauver, J. M., Keelan, J. A., and Mitchell, M. D. (2002). Identification of suppressors of cytokine signaling (SOCS) proteins in human gestational tissues: differential regulation is associated with the onset of labor. J. Clin. Endocrinol. Metab. 87, 1094-1097. - PubMed
-
- Bowen, J. M., Chamley, L., Keelan, J. A., and Mitchell, M. D. (2002). Cytokines of the placenta and extra-placental membranes: roles and regulation during human pregnancy and parturition. Placenta 23, 257-273. - PubMed
-
- Cameron, E. E., Bachman, K. E., Myohanen, S., Herman, J., and Baylin, S. B. (1999). Synergy of demethylation and histone deacetylase inhibition in the re-expression of genes silenced in cancer. Nat. Genet. 21, 103-107. - PubMed
-
- Challis, J.R.G., Matthews, S. G., Gibb, W., and Lye, S. J. (2000). Endocrine and paracrine regulation of birth at term and preterm. Endocr. Rev. 21, 514-550. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
