

Cooperative glutamatergic and cholinergic mechanisms generate short-term modifications of synaptic effectiveness in prepositus hypoglossi neurons
- PMID: 16251437
- PMCID: PMC6725563
- DOI: 10.1523/JNEUROSCI.2061-05.2005
Cooperative glutamatergic and cholinergic mechanisms generate short-term modifications of synaptic effectiveness in prepositus hypoglossi neurons
Abstract
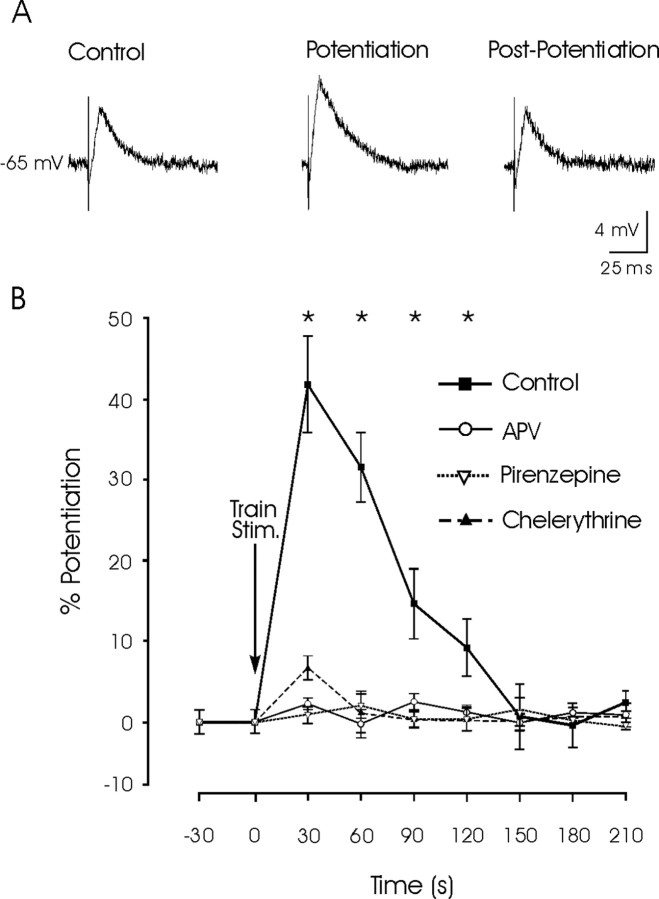
To maintain horizontal eye position on a visual target after a saccade, extraocular motoneurons need a persistent (tonic) neural activity, called "eye-position signal," generated by prepositus hypoglossi (PH) neurons. We have shown previously in vitro and in vivo that this neural activity depends, among others mechanisms, on the interplay of glutamatergic transmission and cholinergic synaptically triggered depolarization. Here, we used rat sagittal brainstem slices, including PH nucleus and paramedian pontine reticular formation (PPRF). We made intracellular recordings of PH neurons and studied their synaptic activation from PPRF neurons. Train stimulation of the PPRF area evoked a cholinergic-sustained depolarization of PH neurons that outlasted the stimulus. EPSPs evoked in PH neurons by single pulses applied to the PPRF presented a short-term potentiation (STP) after train stimulation. APV (an NMDA-receptor blocker) or chelerythrine (a protein kinase-C inhibitor) had no effect on the sustained depolarization, but they did block the evoked STP, whereas pirenzepine (an M1 muscarinic antagonist) blocked both the sustained depolarization and the STP of PH neurons. Thus, electrical stimulation of the PPRF area activates both glutamatergic and cholinergic axons terminating in the PH nucleus, the latter producing a sustained depolarization probably involved in the genesis of the persistent neural activity required for eye fixation. M1-receptor activation seems to evoke a STP of PH neurons via NMDA receptors. Such STP could be needed for the stabilization of the neural network involved in the generation of position signals necessary for eye fixation after a saccade.
Figures



Similar articles
-
A cholinergic synaptically triggered event participates in the generation of persistent activity necessary for eye fixation.J Neurosci. 2004 Jun 2;24(22):5109-18. doi: 10.1523/JNEUROSCI.0235-04.2004. J Neurosci. 2004. PMID: 15175380 Free PMC article.
-
A cholinergic mechanism underlies persistent neural activity necessary for eye fixation.Prog Brain Res. 2006;154:211-24. doi: 10.1016/S0079-6123(06)54011-7. Prog Brain Res. 2006. PMID: 17010712 Review.
-
Cholinergic modulation of primary afferent glutamatergic transmission in rat medullary dorsal horn neurons.Neuropharmacology. 2013 Dec;75:295-303. doi: 10.1016/j.neuropharm.2013.07.030. Epub 2013 Aug 13. Neuropharmacology. 2013. PMID: 23954675
-
Nicotine recruits a local glutamatergic circuit to excite septohippocampal GABAergic neurons.Eur J Neurosci. 2003 Sep;18(5):1155-68. doi: 10.1046/j.1460-9568.2003.02847.x. Eur J Neurosci. 2003. PMID: 12956714
-
GABAB receptors in the medial septum/diagonal band slice from 16-25 day rat.Neuroscience. 2005;132(3):789-800. doi: 10.1016/j.neuroscience.2005.01.027. Neuroscience. 2005. PMID: 15837139
Cited by
-
Nicotinic acetylcholine receptor-mediated responses in medial vestibular and prepositus hypoglossi nuclei neurons showing distinct neurotransmitter phenotypes.J Neurophysiol. 2016 Jun 1;115(5):2649-57. doi: 10.1152/jn.00852.2015. Epub 2016 Mar 2. J Neurophysiol. 2016. PMID: 26936981 Free PMC article.
-
Different Activation Mechanisms of Excitatory Networks in the Rat Oculomotor Integrators for Vertical and Horizontal Gaze Holding.eNeuro. 2020 Jan 21;7(1):ENEURO.0364-19.2019. doi: 10.1523/ENEURO.0364-19.2019. Print 2020 Jan/Feb. eNeuro. 2020. PMID: 31852758 Free PMC article.
-
Neurological basis for eye movements of the blind.PLoS One. 2013;8(2):e56556. doi: 10.1371/journal.pone.0056556. Epub 2013 Feb 18. PLoS One. 2013. PMID: 23441203 Free PMC article.
-
Comparisons of Neuronal and Excitatory Network Properties between the Rat Brainstem Nuclei that Participate in Vertical and Horizontal Gaze Holding.eNeuro. 2017 Sep 13;4(5):ENEURO.0180-17.2017. doi: 10.1523/ENEURO.0180-17.2017. eCollection 2017 Sep-Oct. eNeuro. 2017. PMID: 28966973 Free PMC article.
-
Gaze shift duration, independent of amplitude, influences the number of spikes in the burst for medium-lead burst neurons in pontine reticular formation.Exp Brain Res. 2011 Oct;214(2):225-39. doi: 10.1007/s00221-011-2823-8. Epub 2011 Aug 14. Exp Brain Res. 2011. PMID: 21842410 Free PMC article.
References
-
- Arnold DB, Robinson DA, Leigh RJ (1999) Nystagmus induced by pharmacological inactivation of the brainstem ocular motor integrator in monkey. Vision Res 39: 4286-4295. - PubMed
-
- Calabresi P, Centonze D, Gubellini P, Pisani A, Bernardi G (1998) Endogenous Ach enhances striatal NMDA-responses via M1-like muscarinic receptors and PKC activation. Eur J Neurosci 10: 2887-2895. - PubMed
-
- Carpenter MB, Chang L, Pereira AB, Hersh LB (1987) Comparisons of the immunocytochemical localization of choline acetyltransferase in the vestibular nuclei of the monkey and rat. Brain Res 418: 403-408. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources