

Dysfunction of the cholesterol biosynthetic pathway in Huntington's disease
- PMID: 16251441
- PMCID: PMC6725556
- DOI: 10.1523/JNEUROSCI.3355-05.2005
Dysfunction of the cholesterol biosynthetic pathway in Huntington's disease
Erratum in
-
Erratum: Valenza et al., "Dysfunction of the Cholesterol Biosynthetic Pathway in Huntington's Disease".J Neurosci. 2020 Sep 16;40(38):7375. doi: 10.1523/JNEUROSCI.1842-20.2020. Epub 2020 Sep 1. J Neurosci. 2020. PMID: 32873721 Free PMC article. No abstract available.
Abstract
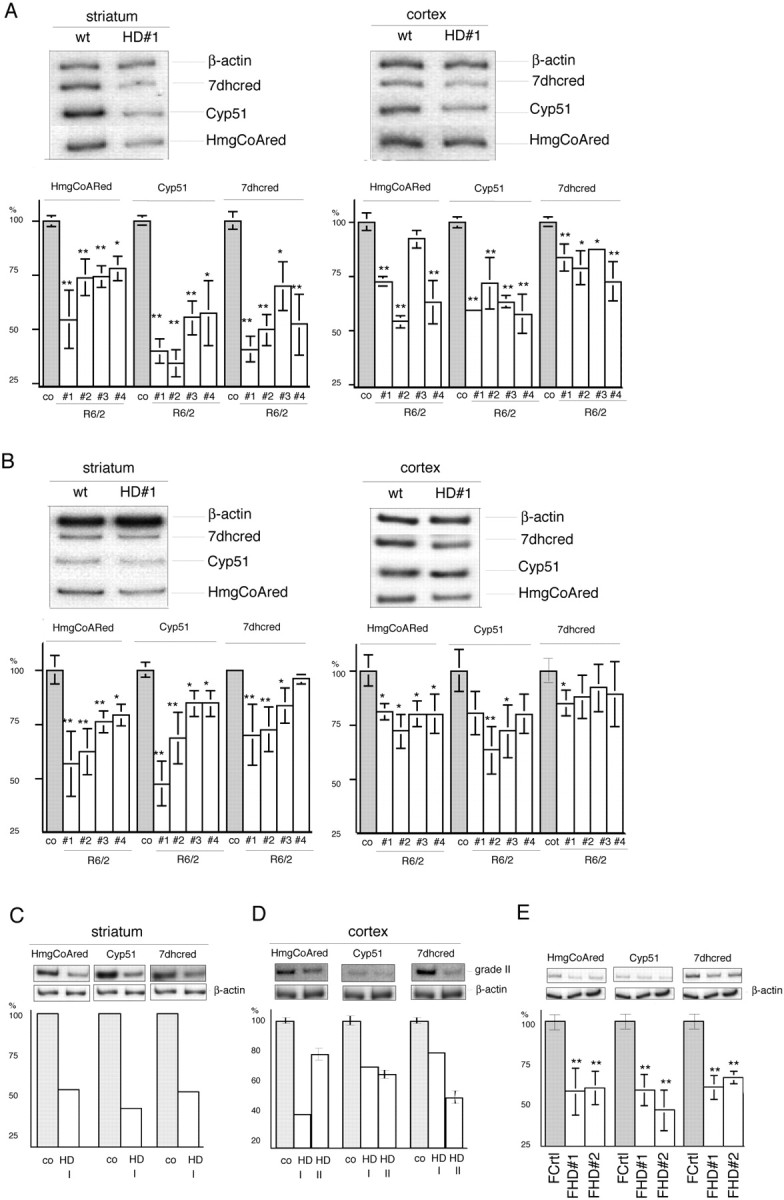
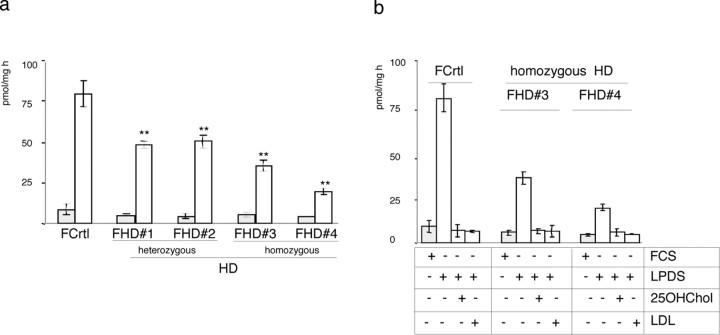
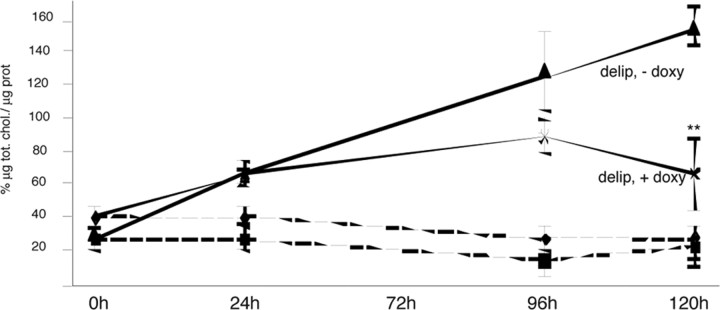
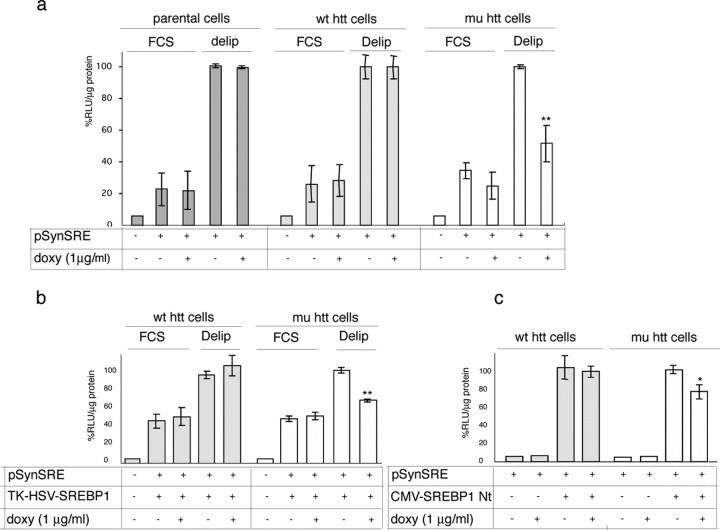
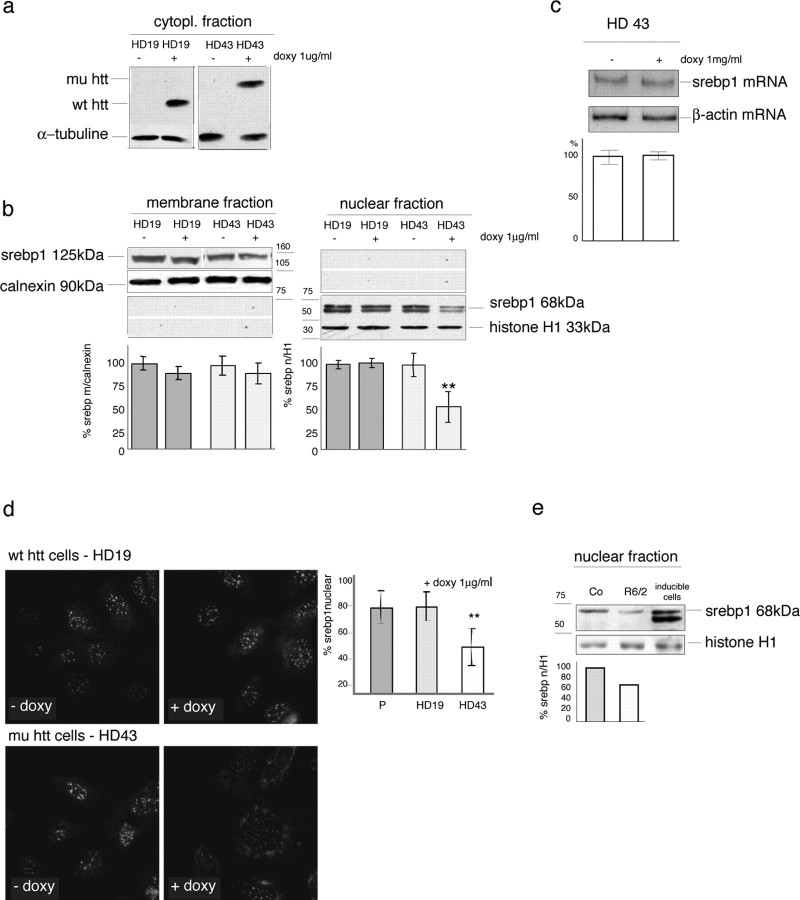
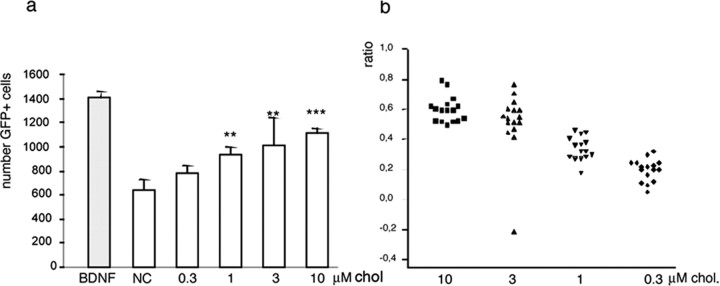
The expansion of a polyglutamine tract in the ubiquitously expressed huntingtin protein causes Huntington's disease (HD), a dominantly inherited neurodegenerative disease. We show that the activity of the cholesterol biosynthetic pathway is altered in HD. In particular, the transcription of key genes of the cholesterol biosynthetic pathway is severely affected in vivo in brain tissue from HD mice and in human postmortem striatal and cortical tissue; this molecular dysfunction is biologically relevant because cholesterol biosynthesis is reduced in cultured human HD cells, and total cholesterol mass is significantly decreased in the CNS of HD mice and in brain-derived ST14A cells in which the expression of mutant huntingtin has been turned on. The transcription of the genes of the cholesterol biosynthetic pathway is regulated via the activity of sterol regulatory element-binding proteins (SREBPs), and we found an approximately 50% reduction in the amount of the active nuclear form of SREBP in HD cells and mouse brain tissue. As a consequence, mutant huntingtin reduces the transactivation of an SRE-luciferase construct even under conditions of SREBP overexpression or in the presence of an exogenous N-terminal active form of SREBP. Finally, the addition of exogenous cholesterol to striatal neurons expressing mutant huntingtin prevents their death in a dose-dependent manner. We conclude that the cholesterol biosynthetic pathway is impaired in HD cells, mice, and human subjects, and that the search for HD therapies should also consider cholesterol levels as both a potential target and disease biomarker.
Figures
References
-
- Barres BA, Smith SJ (2001) Cholesterol—making or breaking the synapse. Science 294: 1296-1297. - PubMed
-
- Battaile KP, Steiner RD (2000) Smith-Lemli-Opitz syndrome: the first mal-formation syndrome associated with defective cholesterol synthesis. Mol Genet Metab 71: 154-162. - PubMed
-
- Bennett MK, Toth JI, Osborne TF (2004) Selective association of sterol regulatory element-binding protein isoforms with target promoters in vivo. J Biol Chem 279: 37360-37367. - PubMed
-
- Brown AJ, Sun L, Feramisco JD, Brown MS, Goldstein JL (2002) Cholesterol addition to ER membranes alters conformation of SCAP, the SREBP escort protein that regulates cholesterol metabolism. Mol Cell 10: 237-245. - PubMed
-
- Brown MS, Goldstein JL (1997) The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 89: 331-340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials