

Restoration of TGF-beta signalling reduces tumorigenicity in human lung cancer cells
- PMID: 16251876
- PMCID: PMC2361493
- DOI: 10.1038/sj.bjc.6602831
Restoration of TGF-beta signalling reduces tumorigenicity in human lung cancer cells
Abstract
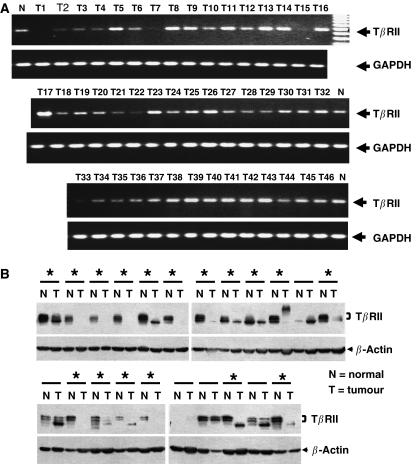
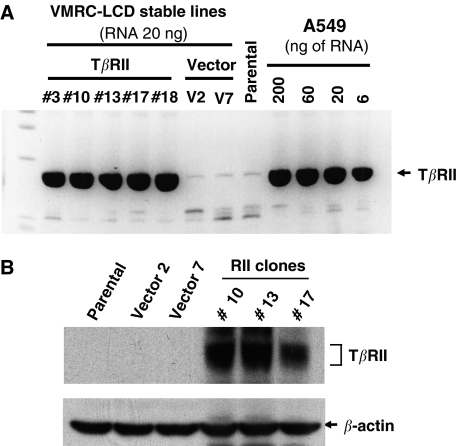
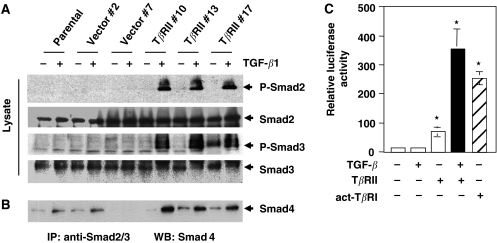
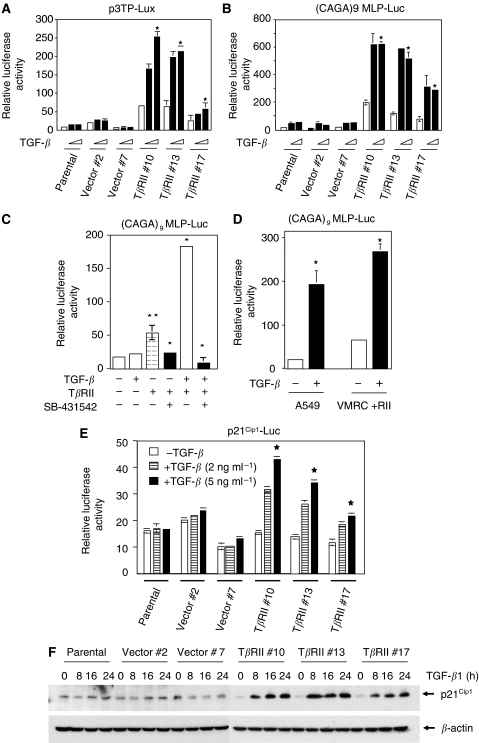
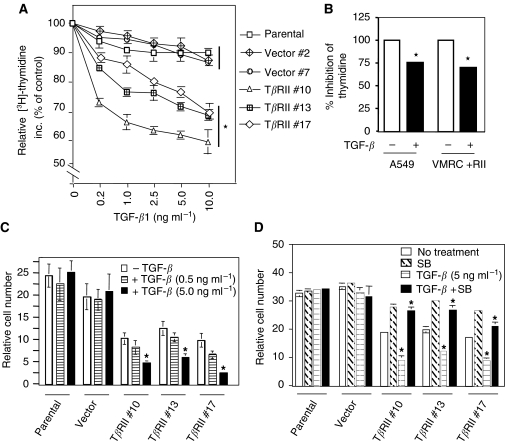
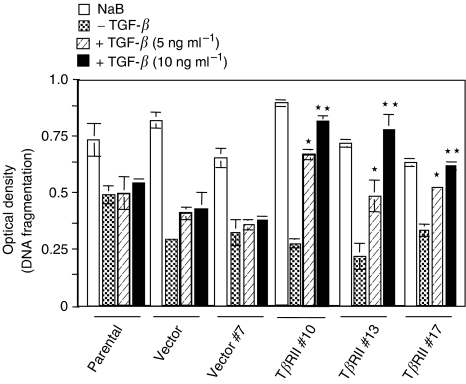
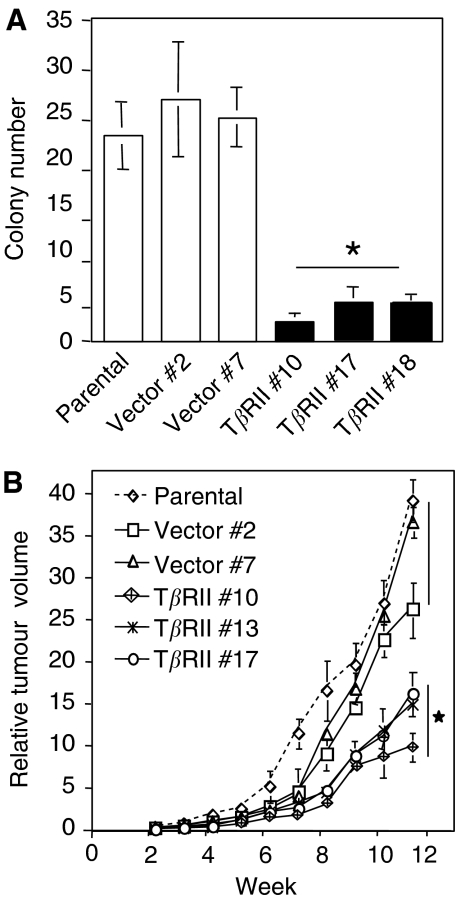
Members of the transforming growth factor-beta (TGF-beta) family regulate a wide range of biological processes including cell proliferation, migration, differentiation, apoptosis, and extracellular matrix deposition. Resistance to TGF-beta-mediated tumour suppressor function in human lung cancer may occur through the loss of type II receptor (TbetaRII) expression. In this study, we investigated the expression pattern of TbetaRII in human lung cancer tissues by RT-PCR and Western blot analyses. We observed downregulation of TbetaRII in 30 out of 46 NSCLC samples (65%) by semiquantitative RT-PCR. Western blot analyses with tumour lysates showed reduced expression of TbetaRII in 77% cases. We also determined the effect of TbetaRII expression in lung adenocarcinoma cell line (VMRC-LCD) that is not responsive to TGF-beta due to lack of TbetaRII expression. Stable expression of TbetaRII in these cells restored TGF-beta-mediated effects including Smad2/3 and Smad4 complex formation, TGF-beta-responsive reporter gene activation, inhibition of cell proliferation and increased apoptosis. Clones expressing TbetaRII showed reduced colony formation in soft-agarose assay and significantly reduced tumorigenicity in athymic nude mice. Therefore, these results suggest that reestablishment of TGF-beta signalling in TbetaRII null cells by stable expression of TbetaRII can reverse malignant behaviour of cells and loss of TbetaRII expression may be involved in lung tumour progression.
Figures
References
-
- Arteaga CL, Tandon AK, VonHoff DD, Osborne CK (1988) Transforming growth factor beta: potential autocrine growth inhibitor of estrogen receptor-negative human breast cancer cells. Cancer Res 48: 3898–3904 - PubMed
-
- Attisano L, Wrana JL (1998) Mads and Smads in TGF beta signalling. Curr Opin Cell Biol 10: 188–194 - PubMed
-
- Datta PK, Chytil A, Gorska AE, Moses HL (1998) Identification of STRAP, a novel WD domain protein in transforming growth factor-beta signaling. J Biol Chem 273: 34671–34674 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
