

Long-QT syndrome-related sodium channel mutations probed by the dynamic action potential clamp technique
- PMID: 16254012
- PMCID: PMC1464302
- DOI: 10.1113/jphysiol.2005.096578
Long-QT syndrome-related sodium channel mutations probed by the dynamic action potential clamp technique
Abstract
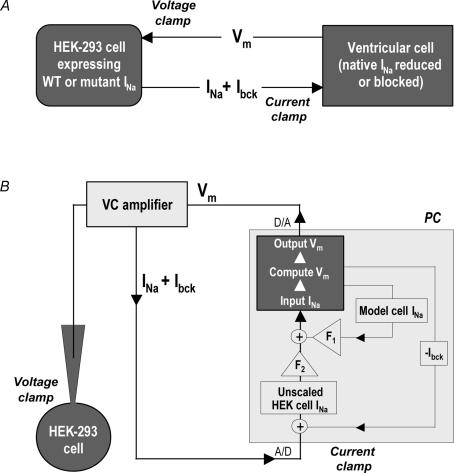
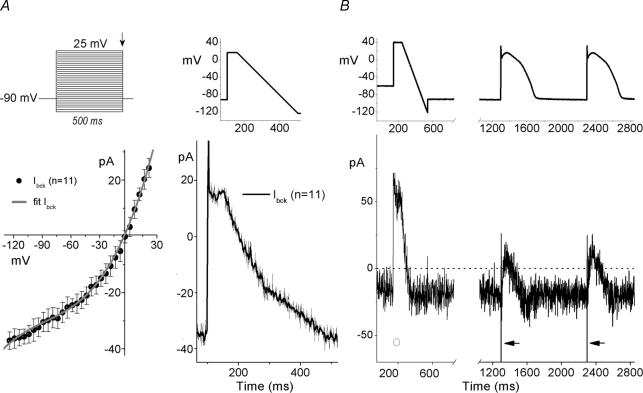
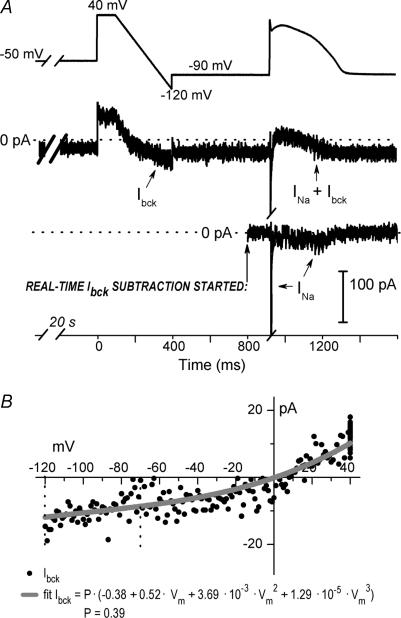
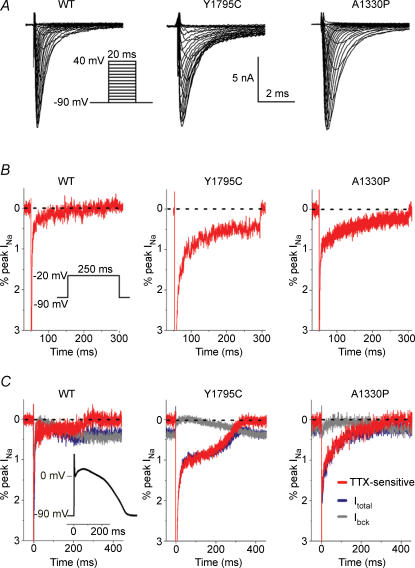
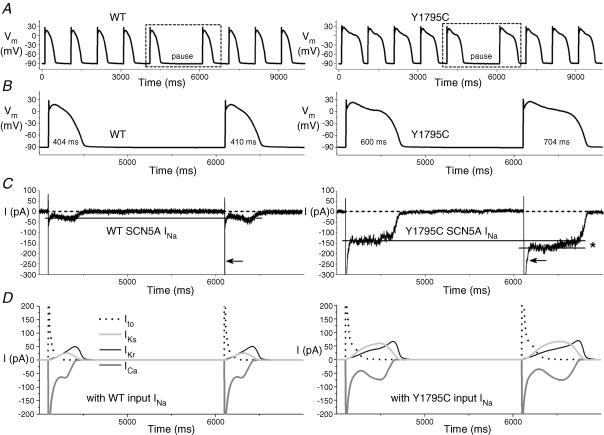
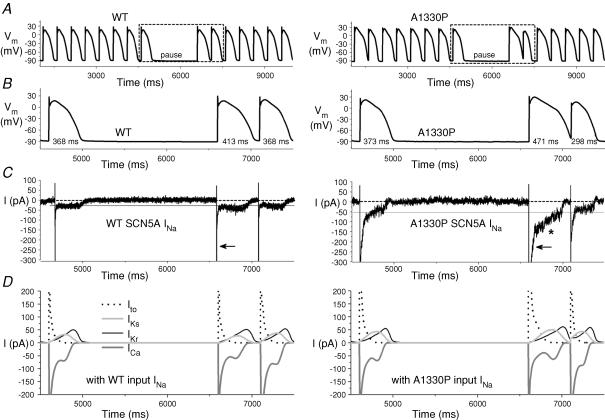
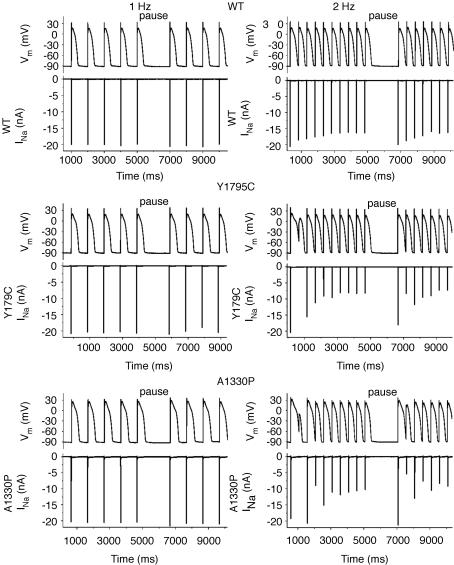
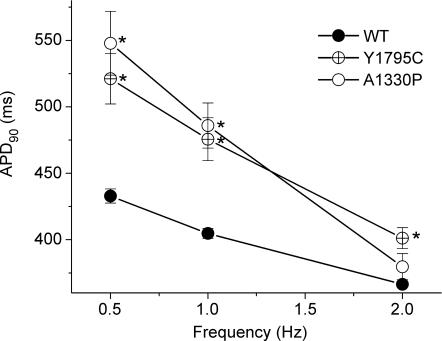
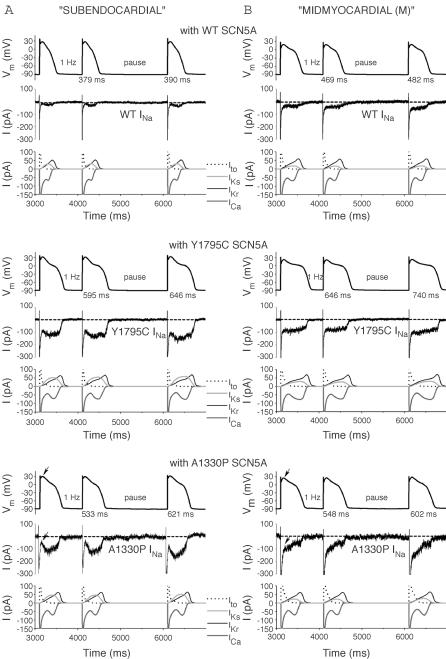
Long-QT3 syndrome (LQT3) is linked to cardiac sodium channel gene (SCN5A) mutations. In this study, we used the 'dynamic action potential clamp' (dAPC) technique to effectively replace the native sodium current (I(Na)) of the Priebe-Beuckelmann human ventricular cell model with wild-type (WT) or mutant I(Na) generated in a human embryonic kidney (HEK)-293 cell that is voltage clamped by the free-running action potential of the ventricular cell. We recorded I(Na) from HEK cells expressing either WT or LQT3-associated Y1795C or A1330P SCN5A at 35 degrees C, and let this current generate and shape the action potential (AP) of subepicardial, mid-myocardial and subendocardial model cells. The HEK cell's endogenous background current was completely removed by a real-time digital subtraction procedure. With WT I(Na), AP duration (APD) was longer than with the original Priebe-Beuckelmann model I(Na), due to a late I(Na) component of approximately 30 pA that could not be revealed with conventional voltage-clamp protocols. With mutant I(Na), this late component was larger ( approximately 100 pA), producing a marked increase in APD ( approximately 70-80 ms at 1 Hz for the subepicardial model cell). The late I(Na) magnitude showed reverse frequency dependence, resulting in a significantly steeper APD-frequency relation in the mutant case. AP prolongation was more pronounced for the mid-myocardial cell type, resulting in increased APD dispersion for each of the mutants. For both mutants, a 2 s pause following rapid (2 Hz) pacing resulted in distorted AP morphology and beat-to-beat fluctuations of I(Na). Our dAPC data directly demonstrate the arrhythmogenic nature of LQT3-associated SCN5A mutations.
Figures
References
-
- An RH, Wang XL, Kerem B, Benhorin J, Medina A, Goldmit M, Kass RS. Novel LQT-3 mutation affects Na+ channel activity through interactions between α- and β1-subunits. Circ Res. 1998;83:141–146. - PubMed
-
- Bennett PB, Yazawa K, Makita N, George AL., Jr Molecular mechanism for an inherited cardiac arrhythmia. Nature. 1995;376:683–685. - PubMed
-
- Carmeliet E. Intracellular Ca2+ concentration and vate adaptatior of the cardiac action potential. Cell Calcium. 2004;35:557–573. - PubMed
-
- Catterall WA. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron. 2000;26:13–25. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous