

Depletion of TDP 43 overrides the need for exonic and intronic splicing enhancers in the human apoA-II gene
- PMID: 16254078
- PMCID: PMC1270946
- DOI: 10.1093/nar/gki897
Depletion of TDP 43 overrides the need for exonic and intronic splicing enhancers in the human apoA-II gene
Erratum in
-
Editor's note on 'Depletion of TDP 43 overrides the need for exonic and intronic splicing enhancers in the human apoA-II gene'.Nucleic Acids Res. 2023 Nov 10;51(20):11411-11413. doi: 10.1093/nar/gkad854. Nucleic Acids Res. 2023. PMID: 37791846 Free PMC article. No abstract available.
Expression of concern in
-
Expression of concern to 'Depletion of TDP 43 overrides the need for exonic and intronic splicing enhancers in the human apoA-II gene'.Nucleic Acids Res. 2023 Feb 28;51(4):2000. doi: 10.1093/nar/gkad113. Nucleic Acids Res. 2023. PMID: 36772831 Free PMC article. No abstract available.
Abstract
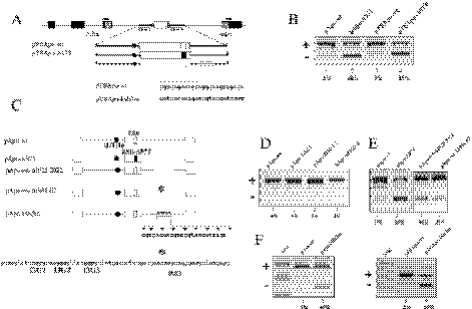
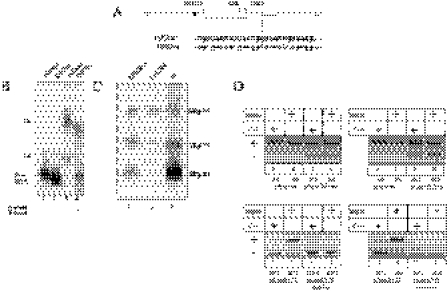
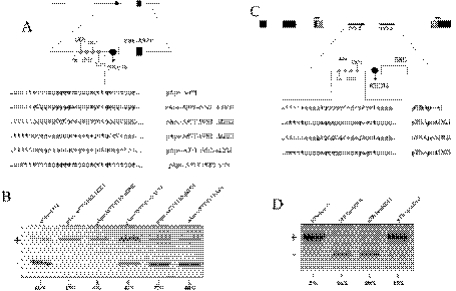
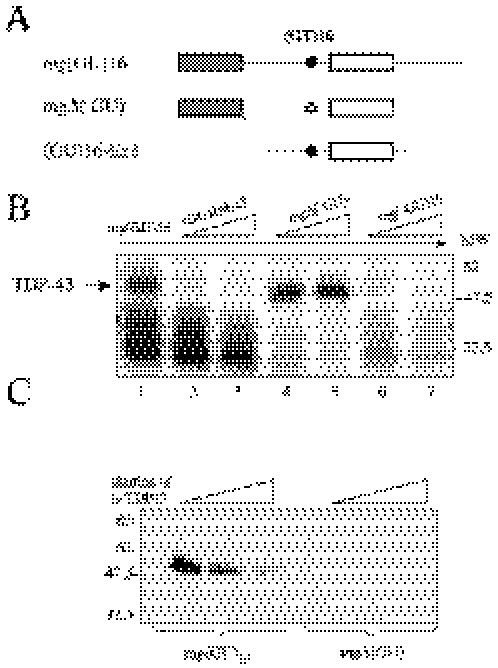
Exon 3 of the human apolipoprotein A-II (apoA-II) gene is efficiently included in the mRNA although its acceptor site is significantly weak because of a peculiar (GU)16 tract instead of a canonical polypyrimidine tract within the intron 2/exon 3 junction. Our previous studies demonstrated that the SR proteins ASF/SF2 and SC35 bind specifically an exonic splicing enhancer (ESE) within exon 3 and promote exon 3 splicing. In the present study, we show that the ESE is necessary only in the proper context. In addition, we have characterized two novel sequences in the flanking introns that modulate apoA-II exon 3 splicing. There is a G-rich element in intron 2 that interacts with hnRNPH1 and inhibits exon 3 splicing. The second is a purine rich region in intron 3 that binds SRp40 and SRp55 and promotes exon 3 inclusion in mRNA. We have also found that the (GU) repeats in the apoA-II context bind the splicing factor TDP-43 and interfere with exon 3 definition. Significantly, blocking of TDP-43 expression by small interfering RNA overrides the need for all the other cis-acting elements making exon 3 inclusion constitutive even in the presence of disrupted exonic and intronic enhancers. Altogether, our results suggest that exonic and intronic enhancers have evolved to balance the negative effects of the two silencers located in intron 2 and hence rescue the constitutive exon 3 inclusion in apoA-II mRNA.
Figures

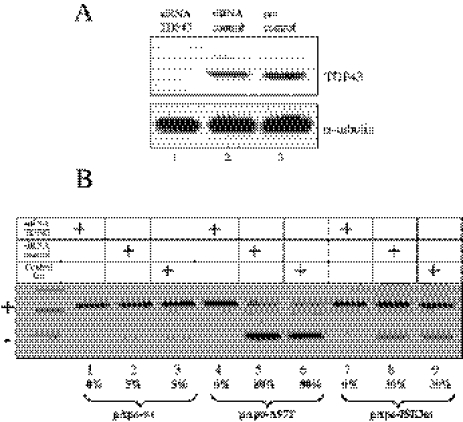
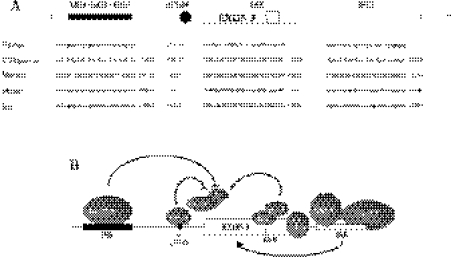
Comment in
-
Editor's note on 'Depletion of TDP 43 overrides the need for exonic and intronic splicing enhancers in the human apoA-II gene'.Nucleic Acids Res. 2023 Nov 10;51(20):11411-11413. doi: 10.1093/nar/gkad854. Nucleic Acids Res. 2023. PMID: 37791846 Free PMC article. No abstract available.
References
-
- Smith C.W., Valcarcel J. Alternative pre-mRNA splicing: the logic of combinatorial control. Trends Biochem. Sci. 2000;25:381–388. - PubMed
-
- Black D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003;72:291–336. - PubMed
-
- Blencowe B.J. Exonic splicing enhancers: mechanism of action, diversity and role in human genetic diseases. Trends Biochem. Sci. 2000;25:106–110. - PubMed
-
- Hovhannisyan R.H., Carstens R.P. A novel intronic cis element, ISE/ISS-3, regulates rat fibroblast growth factor receptor 2 splicing through activation of an upstream exon and repression of a downstream exon containing a noncanonical branch point sequence. Mol. Cell. Biol. 2005;25:250–263. - PMC - PubMed
-
- Arrisi-Mercado P., Romano M., Muro A.F., Baralle F.E. An exonic splicing enhancer offsets the atypical GU-rich 3′ splice site of human apolipoprotein A-II exon 3. J. Biol. Chem. 2004;279:39331–39339. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
