

Transcriptional regulation of early transposon elements, an active family of mouse long terminal repeat retrotransposons
- PMID: 16254322
- PMCID: PMC1280189
- DOI: 10.1128/JVI.79.22.13865-13874.2005
Transcriptional regulation of early transposon elements, an active family of mouse long terminal repeat retrotransposons
Abstract
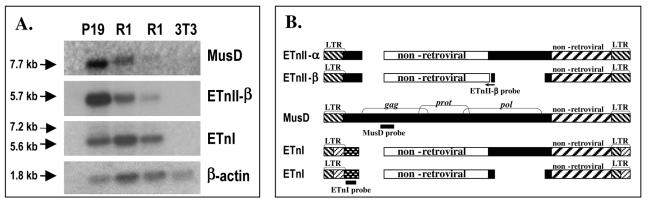
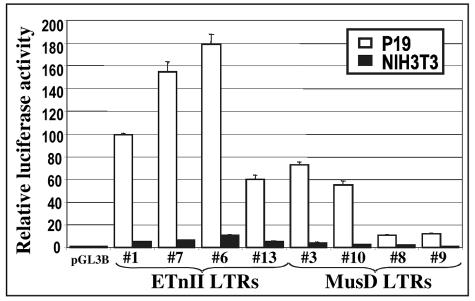
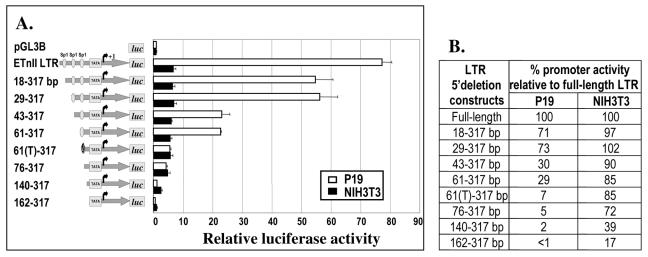
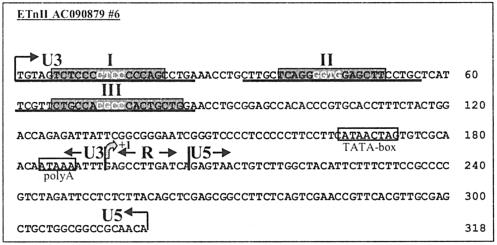
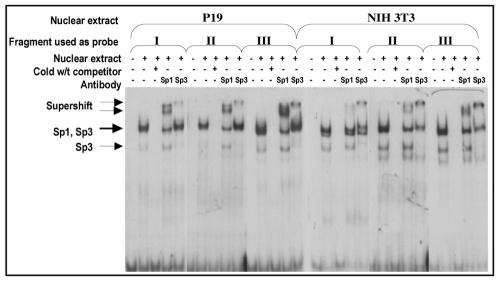
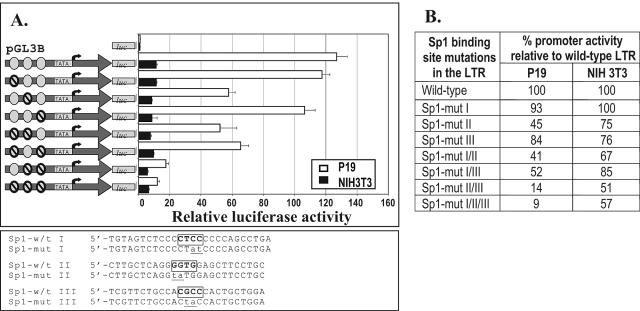
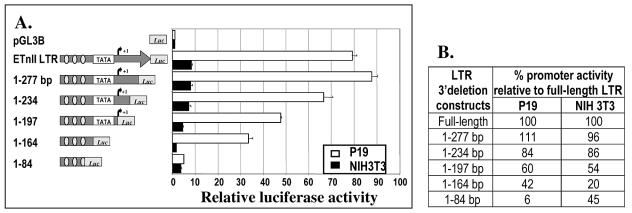
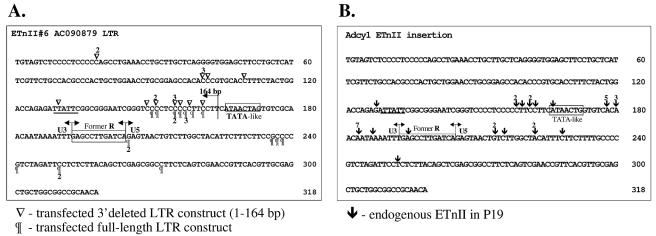
While early transposon (ETn) endogenous retrovirus (ERV)-like elements are known to be active insertional mutagens in the mouse, little is known about their transcriptional regulation. ETns are transcribed during early mouse embryogenesis in embryonic stem (ES) and embryonic carcinoma (EC) cell lines. Despite their lack of coding potential, some ETns remain transposition competent through their use of reverse transcriptase encoded by a related group of ERVs-MusD elements. In this study, we have confirmed high expression levels of ETn and MusD elements in ES and EC cells and have demonstrated an increase in the copy number of ETnII elements in the EC P19 cell line. Using transient transfections, we have shown that ETnII and MusD LTRs are much more active as promoters in P19 cells than in NIH 3T3 cells, indicating that genomic context and methylation are not the only factors determining endogenous transcriptional activity of ETns. Three sites in the 5' part of the long terminal repeat (LTR) were demonstrated to bind Sp1 and Sp3 transcription factors and were found to be important for high LTR promoter activity in P19 cells, suggesting that as yet unidentified Sp binding partners are involved in the regulation of ETn activity in undifferentiated cells. Finally, we found multiple transcription start sites within the ETn LTR and have shown that the LTR retains significant promoter activity in the absence of its noncanonical TATA box. These findings lend insight into the transcriptional regulation of this family of mobile mouse retrotransposons.
Figures
Similar articles
-
An active murine transposon family pair: retrotransposition of "master" MusD copies and ETn trans-mobilization.Genome Res. 2004 Nov;14(11):2261-7. doi: 10.1101/gr.2924904. Epub 2004 Oct 12. Genome Res. 2004. PMID: 15479948 Free PMC article.
-
Novel mouse type D endogenous proviruses and ETn elements share long terminal repeat and internal sequences.J Virol. 2000 Aug;74(16):7221-9. doi: 10.1128/jvi.74.16.7221-7229.2000. J Virol. 2000. PMID: 10906176 Free PMC article.
-
Endogenous retrovirus long terminal repeats as ready-to-use mobile promoters: the case of primate beta3GAL-T5.Gene. 2005 Dec 30;364:2-12. doi: 10.1016/j.gene.2005.05.045. Epub 2005 Aug 22. Gene. 2005. PMID: 16112824
-
[Retroviruses-derived sequences in the human genome. Human endogenous retroviruses (HERVs)].Postepy Hig Med Dosw (Online). 2006;60:637-52. Postepy Hig Med Dosw (Online). 2006. PMID: 17199106 Review. Polish.
-
Endogenous Retroviruses Walk a Fine Line between Priming and Silencing.Viruses. 2020 Jul 23;12(8):792. doi: 10.3390/v12080792. Viruses. 2020. PMID: 32718022 Free PMC article. Review.
Cited by
-
Functional regulation of an ancestral RAG transposon ProtoRAG by a trans-acting factor YY1 in lancelet.Nat Commun. 2020 Sep 9;11(1):4515. doi: 10.1038/s41467-020-18261-7. Nat Commun. 2020. PMID: 32908127 Free PMC article.
-
Identification and promoter analysis of PERV LTR subtypes in NIH-miniature pig.Mol Cells. 2013 Feb;35(2):99-105. doi: 10.1007/s10059-013-2289-6. Epub 2013 Feb 21. Mol Cells. 2013. PMID: 23456331 Free PMC article.
-
Proviral silencing in embryonic stem cells requires the histone methyltransferase ESET.Nature. 2010 Apr 8;464(7290):927-31. doi: 10.1038/nature08858. Epub 2010 Feb 17. Nature. 2010. PMID: 20164836
-
A reverse transcriptase-dependent mechanism is essential for murine preimplantation development.Genes (Basel). 2011 May 18;2(2):360-73. doi: 10.3390/genes2020360. Genes (Basel). 2011. PMID: 24710196 Free PMC article.
-
Quadruplex-forming sequences occupy discrete regions inside plant LTR retrotransposons.Nucleic Acids Res. 2014 Jan;42(2):968-78. doi: 10.1093/nar/gkt893. Epub 2013 Oct 7. Nucleic Acids Res. 2014. PMID: 24106085 Free PMC article.
References
-
- Asch, B. B., H. L. Asch, D. L. Stoler, and G. R. Anderson. 1993. De-regulation of endogenous retrotransposons in mouse mammary carcinomas of diverse etiologies. Int. J. Cancer 54:813-819. - PubMed
-
- Baust, C., G. J. Baillie, and D. L. Mager. 2002. Insertional polymorphisms of ETn retrotransposons include a disruption of the wiz gene in C57BL/6 mice. Mamm. Genome 13:423-428. - PubMed
-
- Bevilacqua, A., M. T. Fiorenza, and F. Mangia. 2000. A developmentally regulated GAGA box-binding factor and Sp1 are required for transcription of the hsp70.1 gene at the onset of mouse zygotic genome activation. Development 127:1541-1551. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
