

A live attenuated vaccine for Lassa fever made by reassortment of Lassa and Mopeia viruses
- PMID: 16254329
- PMCID: PMC1280243
- DOI: 10.1128/JVI.79.22.13934-13942.2005
A live attenuated vaccine for Lassa fever made by reassortment of Lassa and Mopeia viruses
Abstract
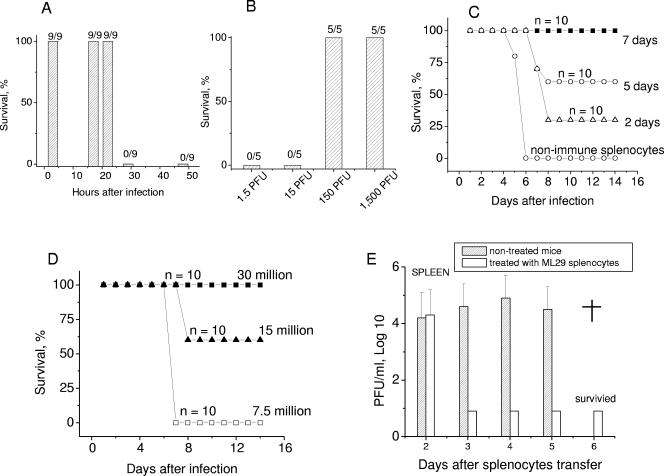
Lassa virus (LASV) and Mopeia virus (MOPV) are closely related Old World arenaviruses that can exchange genomic segments (reassort) during coinfection. Clone ML29, selected from a library of MOPV/LASV (MOP/LAS) reassortants, encodes the major antigens (nucleocapsid and glycoprotein) of LASV and the RNA polymerase and zinc-binding protein of MOPV. Replication of ML29 was attenuated in guinea pigs and nonhuman primates. In murine adoptive-transfer experiments, as little as 150 PFU of ML29 induced protective cell-mediated immunity. All strain 13 guinea pigs vaccinated with clone ML29 survived at least 70 days after LASV challenge without either disease signs or histological lesions. Rhesus macaques inoculated with clone ML29 developed primary virus-specific T cells capable of secreting gamma interferon in response to homologous MOP/LAS and heterologous MOPV and lymphocytic choriomeningitis virus. Detailed examination of two rhesus macaques infected with this MOPV/LAS reassortant revealed no histological lesions or disease signs. Thus, ML29 is a promising attenuated vaccine candidate for Lassa fever.
Figures
References
-
- Anonymous. 2003. Influenza virus vaccine live intranasal-MedImmune vaccines: CAIV-T, influenza vaccine live intranasal. Drugs R&D 4:312-319. - PubMed
-
- Auperin, D. D., D. R. Sasso, and J. B. McCormick. 1986. Nucleotide sequence of the glycoprotein gene and intergenic region of the Lassa virus S genome RNA. Virology 154:155-167. - PubMed
-
- Borio, L., T. Inglesby, C. J. Peters, A. L. Schmaljohn, J. M. Hughes, P. B. Jahrling, T. Ksiazek, K. M. Johnson, A. Meyerhoff, T. O'Toole, M. S. Ascher, J. Bartlett, J. G. Breman, E. M. Eitzen, Jr., M. Hamburg, J. Hauer, D. A. Henderson, R. T. Johnson, G. Kwik, M. Layton, S. Lillibridge, G. J. Nabel, M. T. Osterholm, T. M. Perl, P. Russell, K. Tonat, and the Working Group on Civilian Biodefense. 2002. Hemorrhagic fever viruses as biological weapons: medical and public health management. JAMA 287:2391-2405. - PubMed
-
- Chandler, L. J., G. Hogge, M. Endres, D. R. Jacoby, N. Nathanson, and B. J. Beaty. 1991. Reassortment of La Crosse and Tahyna bunyaviruses in Aedes triseriatus mosquitoes. Virus Res. 20:181-191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
