

Alpha/beta interferons regulate murine gammaherpesvirus latent gene expression and reactivation from latency
- PMID: 16254350
- PMCID: PMC1280204
- DOI: 10.1128/JVI.79.22.14149-14160.2005
Alpha/beta interferons regulate murine gammaherpesvirus latent gene expression and reactivation from latency
Abstract
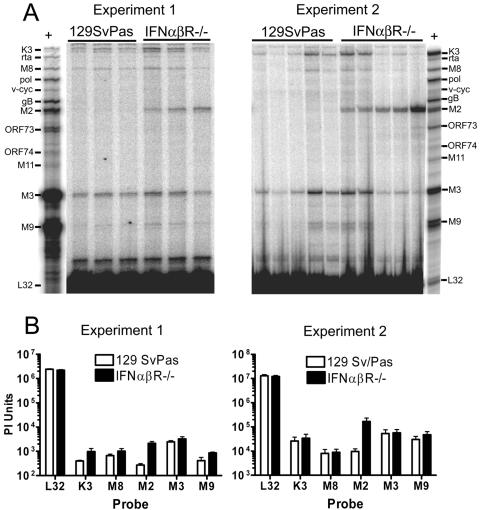
Alpha/beta interferon (IFN-alpha/beta) protects the host from virus infection by inhibition of lytic virus replication in infected cells and modulation of the antiviral cell-mediated immune response. To determine whether IFN-alpha/beta also modulates the virus-host interaction during latent virus infection, we infected mice lacking the IFN-alpha/beta receptor (IFN-alpha/betaR(-/-)) and wild-type (wt; 129S2/SvPas) mice with murine gammaherpesvirus 68 (gammaHV68), a lymphotropic gamma-2-herpesvirus that establishes latent infection in B cells, macrophages, and dendritic cells. IFN-alpha/betaR(-/-) mice cleared low-dose intranasal gammaHV68 infection with wt kinetics and harbored essentially wt frequencies of latently infected cells in both peritoneum and spleen by 28 days postinfection. However, latent virus in peritoneal cells and splenocytes from IFN-alpha/betaR(-/-) mice reactivated ex vivo with >40-fold- and 5-fold-enhanced efficiency, respectively, compared to wt cells. Depletion of IFN-alpha/beta from wt mice during viral latency also significantly increased viral reactivation, demonstrating an antiviral function of IFN-alpha/beta during latency. Viral reactivation efficiency was temporally regulated in both wt and IFN-alpha/betaR(-/-) mice. The mechanism of IFN-alpha/betaR action was distinct from that of IFN-gammaR, since IFN-alpha/betaR(-/-) mice did not display persistent virus replication in vivo. Analysis of viral latent gene expression in vivo demonstrated specific upregulation of the latency-associated gene M2, which is required for efficient reactivation from latency, in IFN-alpha/betaR(-/-) splenocytes. These data demonstrate that an IFN-alpha/beta-induced pathway regulates gammaHV68 gene expression patterns during latent viral infection in vivo and that IFN-alpha/beta plays a critical role in inhibiting viral reactivation during latency.
Figures






References
-
- Agnello, D., C. S. R. Lankford, J. Bream, A. Morinobu, M. Gadina, J. J. O'Shea, and D. M. Frucht. 2003. Cytokines and transcription factors that regulate T helper cell differentiation: new players and new insights. J. Clin. Immunol. 23:147-161. - PubMed
-
- Arico, E., K. A. Robertson, F. Belardelli, M. Ferrantini, and A. A. Nash. 2004. Vaccination with inactivated murine gammaherpesvirus 68 strongly limits viral replication and latency and protects type I IFN receptor knockout mice from a lethal infection. Vaccine 22:1433-1440. - PubMed
-
- Barnes, B., B. Lubyova, and P. M. Pitha. 2002. On the role of IRF in host defense. J. Interferon Cytokine Res. 22:59-71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
