

Protein synthesis required for long-term memory is induced by PKC activation on days before associative learning
- PMID: 16258064
- PMCID: PMC1283453
- DOI: 10.1073/pnas.0508001102
Protein synthesis required for long-term memory is induced by PKC activation on days before associative learning
Abstract
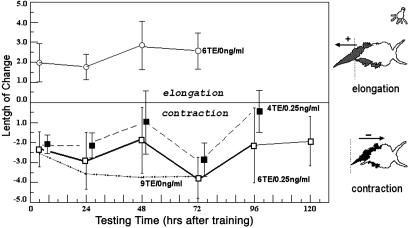
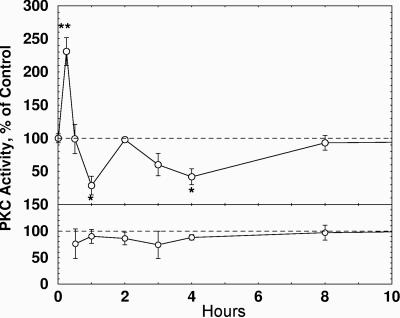
Protein synthesis has long been known to be required for associative learning to consolidate into long-term memory. Here we demonstrate that PKC isozyme activation on days before training can induce the synthesis of proteins necessary and sufficient for subsequent long-term memory consolidation. Bryostatin (Bryo), a macrolide lactone with efficacy in subnanomolar concentrations and a potential therapeutic for Alzheimer's disease, is a potent activator of PKC, some of whose isozymes undergo prolonged activation after associative learning. Under normal conditions, two training events with paired visual and vestibular stimuli cause short-term memory of the mollusc Hermissenda that lasts approximately 7 min. However, after 4-h exposures to Bryo (0.25 ng/ml) on two preceding days, the same two training events produced long-term conditioning that lasted >1 week and that was not blocked by anisomycin (1 mug/ml). Anisomycin, however, eliminated long-term memory lasting at least 1 week after nine training events. Both the nine training events alone and two Bryo exposures plus two training event regimens caused comparably increased levels of the PKC alpha-isozyme substrate calexcitin in identified type B neurons and enhanced PKC activity in the membrane fractions. Furthermore, Bryo increased overall protein synthesis in cultured mammalian neurons by up to 60% for >3 days. The specific PKC antagonist Ro-32-0432 blocked much of this Bryo-induced protein synthesis as well as the Bryo-induced enhancement of the behavioral conditioning. Thus, Bryo-induced PKC activation produces those proteins necessary and sufficient for long-term memory on days in advance of the training events themselves.
Figures









Similar articles
-
Bryostatin enhancement of memory in Hermissenda.Biol Bull. 2006 Jun;210(3):201-14. doi: 10.2307/4134558. Biol Bull. 2006. PMID: 16801495
-
Inhibition of phorbol ester-induced T cell proliferation by bryostatin is associated with rapid degradation of protein kinase C.J Immunol. 1993 Feb 15;150(4):1195-204. J Immunol. 1993. PMID: 8432975
-
Protein kinase C mediates memory consolidation of taste avoidance conditioning in Lymnaea stagnalis.Neurobiol Learn Mem. 2014 May;111:9-18. doi: 10.1016/j.nlm.2014.02.011. Epub 2014 Mar 6. Neurobiol Learn Mem. 2014. PMID: 24613854
-
Synthesis and biological activities of simplified analogs of the natural PKC ligands, bryostatin-1 and aplysiatoxin.Chem Rec. 2014 Apr;14(2):251-67. doi: 10.1002/tcr.201300036. Epub 2014 Feb 20. Chem Rec. 2014. PMID: 24677503 Review.
-
Challenges to the development of bryostatin-type anticancer drugs based on the activation mechanism of protein kinase Cδ.Med Res Rev. 2012 May;32(3):518-35. doi: 10.1002/med.20220. Epub 2010 Nov 9. Med Res Rev. 2012. PMID: 22539107 Review.
Cited by
-
Neural stem cell transplants improve cognitive function without altering amyloid pathology in an APP/PS1 double transgenic model of Alzheimer's disease.Mol Neurobiol. 2014 Oct;50(2):423-37. doi: 10.1007/s12035-014-8640-x. Epub 2014 Jan 31. Mol Neurobiol. 2014. PMID: 24481678
-
Programming of neurotoxic cofactor CXCL-10 in HIV-1-associated dementia: abrogation of CXCL-10-induced neuro-glial toxicity in vitro by PKC activator.J Neuroinflammation. 2012 Oct 18;9:239. doi: 10.1186/1742-2094-9-239. J Neuroinflammation. 2012. PMID: 23078780 Free PMC article.
-
nPKCε Mediates SNAP-25 Phosphorylation of Ser-187 in Basal Conditions and After Synaptic Activity at the Neuromuscular Junction.Mol Neurobiol. 2019 Aug;56(8):5346-5364. doi: 10.1007/s12035-018-1462-5. Epub 2019 Jan 3. Mol Neurobiol. 2019. PMID: 30607888
-
Temporal-Specific Sex and Injury-Dependent Changes on Neurogranin-Associated Synaptic Signaling After Controlled Cortical Impact in Rats.Mol Neurobiol. 2024 Sep;61(9):7256-7268. doi: 10.1007/s12035-024-04043-5. Epub 2024 Feb 20. Mol Neurobiol. 2024. PMID: 38376763
-
Total synthesis of bryostatin 7 via C-C bond-forming hydrogenation.J Am Chem Soc. 2011 Sep 7;133(35):13876-9. doi: 10.1021/ja205673e. Epub 2011 Aug 11. J Am Chem Soc. 2011. PMID: 21780806 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
