

The POU protein Oct-6 is a nucleocytoplasmic shuttling protein
- PMID: 16260476
- PMCID: PMC1275591
- DOI: 10.1093/nar/gki947
The POU protein Oct-6 is a nucleocytoplasmic shuttling protein
Abstract
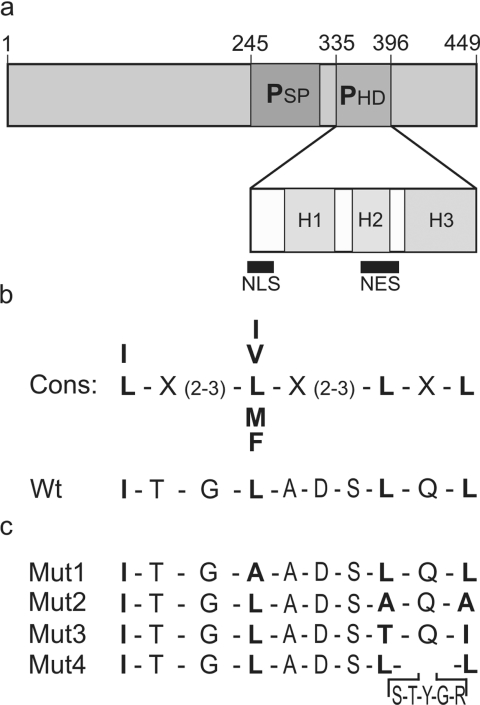
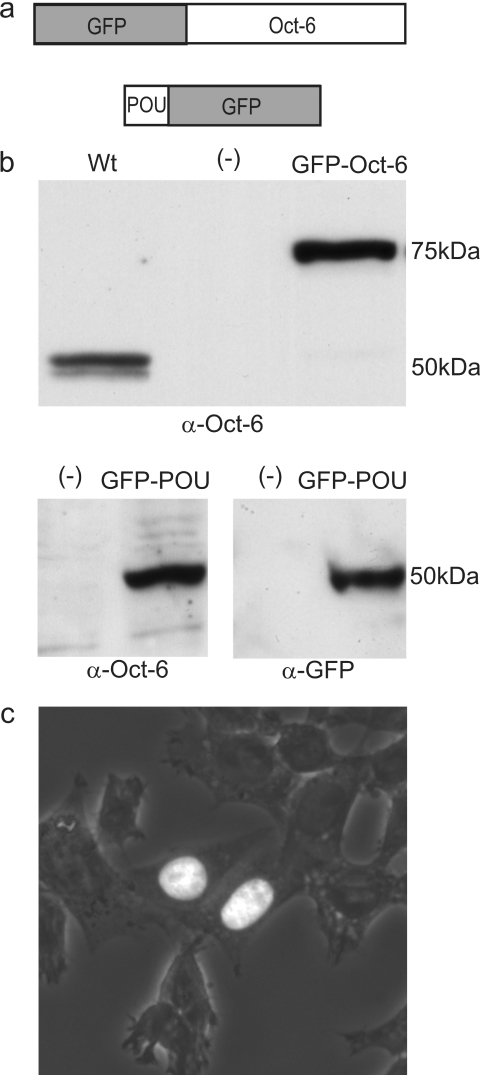
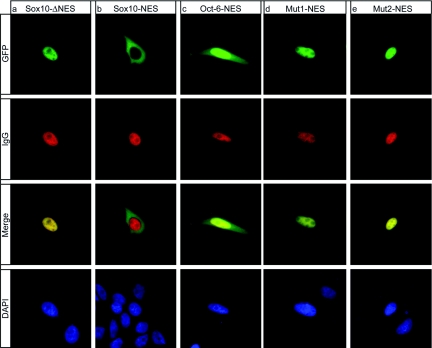
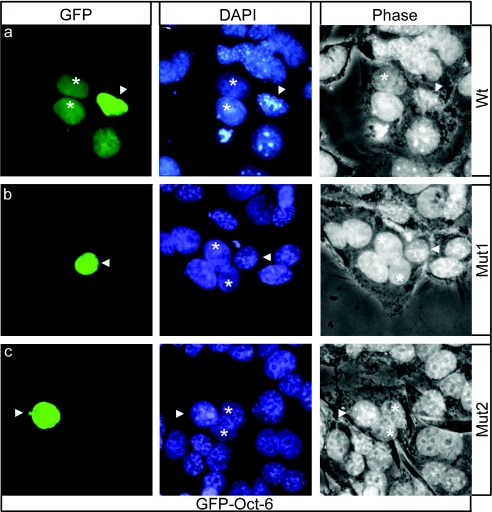
Like many POU domain proteins, Oct-6 plays important roles during vertebrate development. In accord with its function as a transcriptional regulator during neurogenesis and myelination, Oct-6 is predominantly found in the nucleus. Nuclear import is mediated by a nuclear localization signal at the N-terminal end of the POU homeodomain. Here we show, that Oct-6 in addition contains a nuclear export signal so that Oct-6 is able to shuttle constantly between nucleus and cytoplasm. This nuclear export signal is also localized in the POU homeodomain as part of helix 2 and the connecting loop to DNA recognition helix 3. It conforms to the consensus of hydrophobic leucine-rich export sequences and mediates export from the nucleus via CRM1/Exp1. Several amino acid substitutions or insertions that inactivate this nuclear export sequence, reduce DNA-binding of Oct-6 to its octamer recognition element slighty, but interfere strongly with Oct-6-dependent transcriptional activation, thus arguing that nuclear export and nucleocytoplasmic shuttling are essential aspects of Oct-6 function. Importantly, the nuclear export signal identified for Oct-6 is conserved in most, if not all other vertebrate POU proteins. Nuclear export might therefore be of general relevance for POU protein function throughout development.
Figures





References
-
- Ryan A.K., Rosenfeld M.G. POU domain family values: flexibility, partnerships, and developmental codes. Genes Dev. 1997;11:1207–1225. - PubMed
-
- Wegner M., Drolet D.W., Rosenfeld M.G. POU-domain proteins: structure and function of developmental regulators. Curr. Opin. Cell Biol. 1993;5:488–498. - PubMed
-
- Alvarez-Bolado G., Rosenfeld M.G., Swanson L.W. Model of forebrain regionalization based on spatiotemporal patterns of POU-III homeobox gene expression, birthdates, and morphological features. J. Comp. Neurol. 1995;355:237–295. - PubMed
-
- Bermingham J.R., Jr, Scherer S.S., O'Connell S., Arroyo E., Kalla K.A., Powell F.L., Rosenfeld M.G. Tst-1/Oct-6/SCIP regulates a unique step in peripheral myelination and is required for normal respiration. Genes Dev. 1996;10:1751–1762. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
