

The insulin A-chain epitope recognized by human T cells is posttranslationally modified
- PMID: 16260488
- PMCID: PMC2213236
- DOI: 10.1084/jem.20051251
The insulin A-chain epitope recognized by human T cells is posttranslationally modified
Abstract
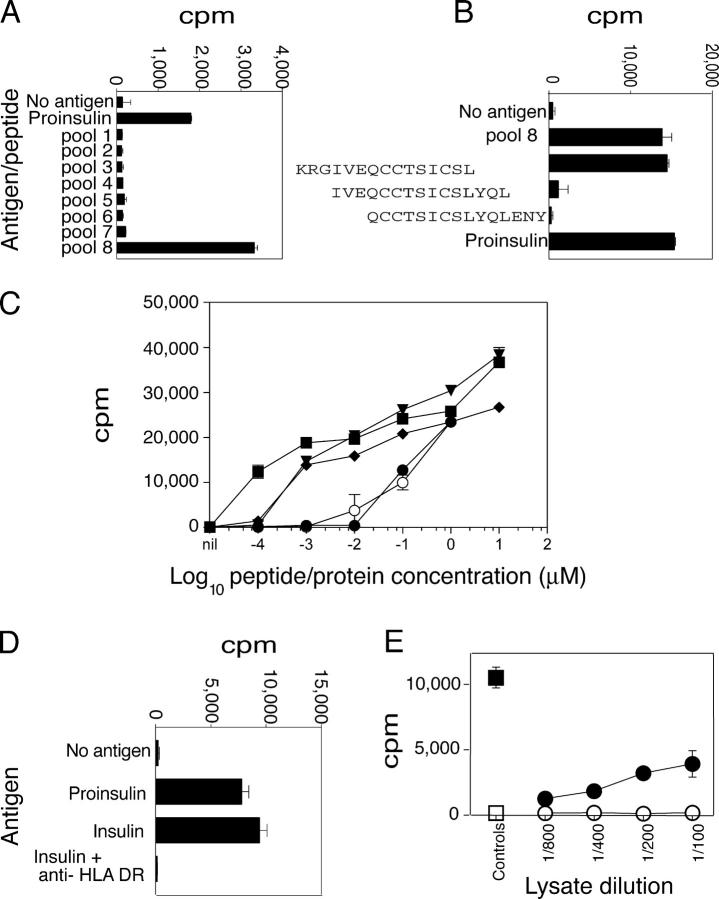
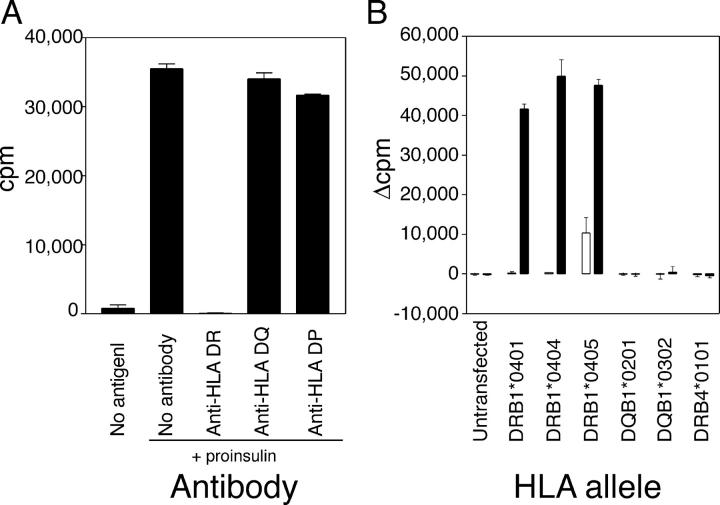
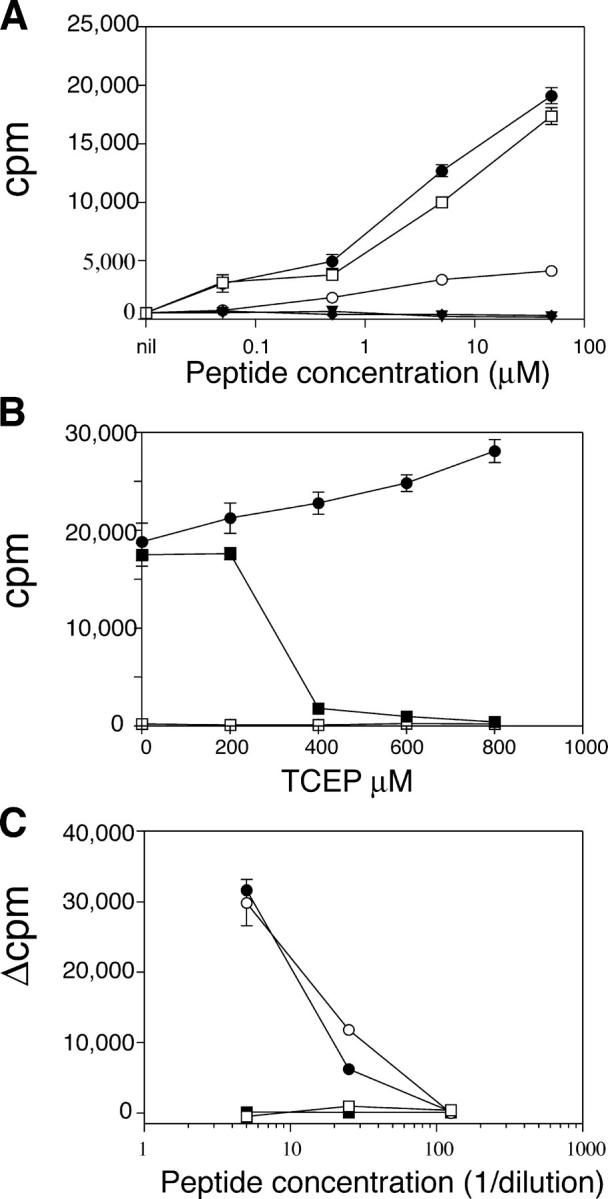
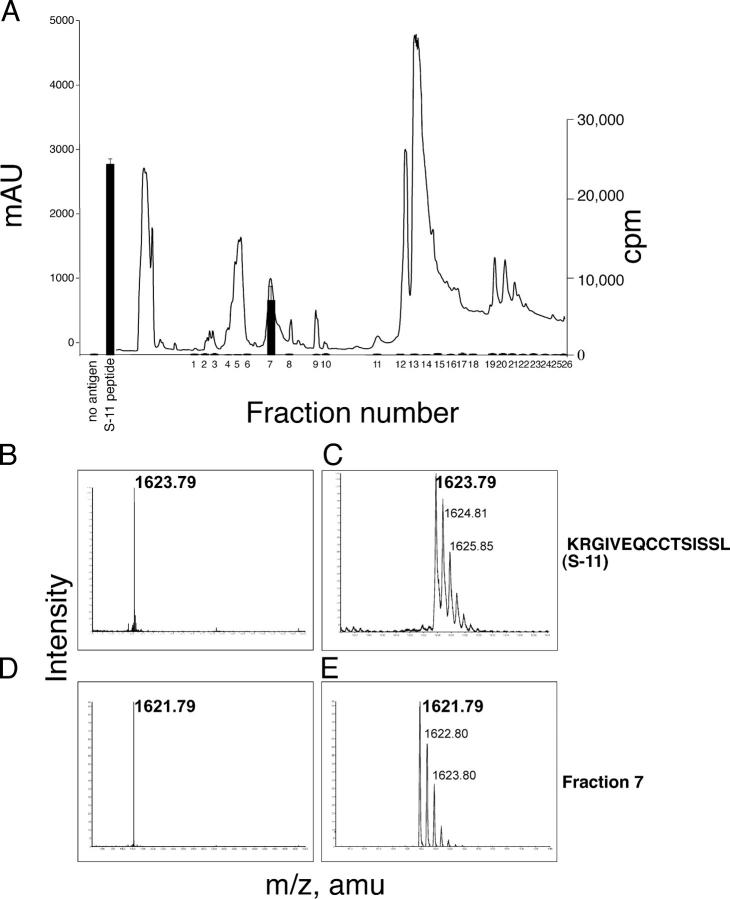
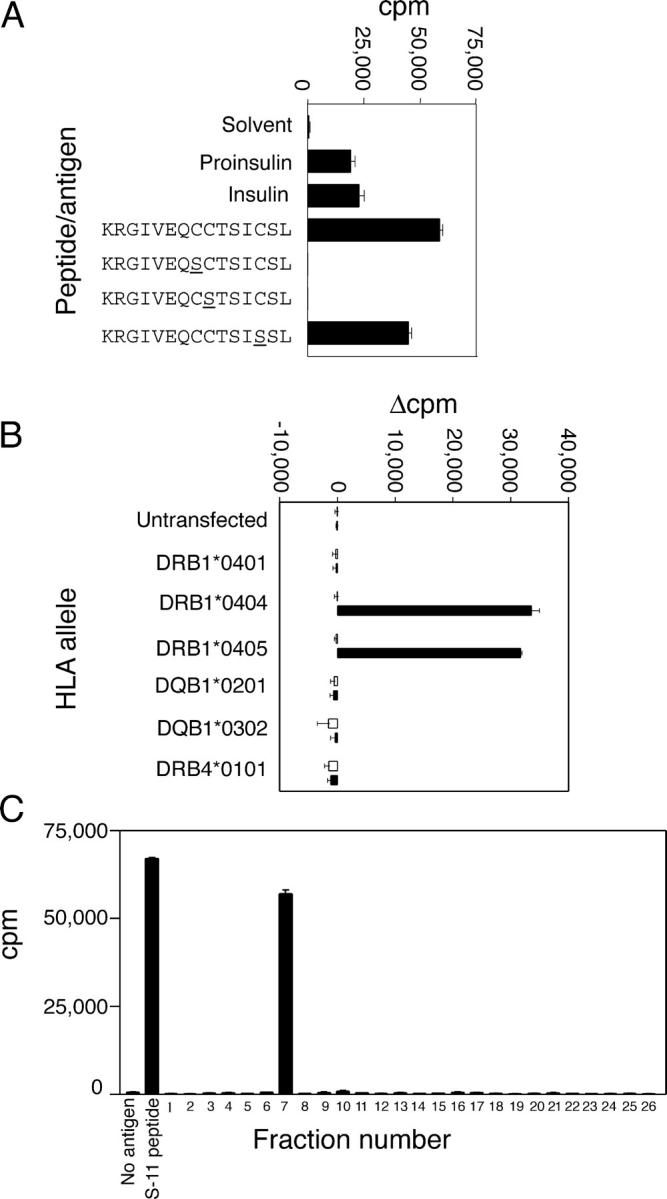
The autoimmune process that destroys the insulin-producing pancreatic beta cells in type 1 diabetes (T1D) is targeted at insulin and its precursor, proinsulin. T cells that recognize the proximal A-chain of human insulin were identified recently in the pancreatic lymph nodes of subjects who had T1D. To investigate the specificity of proinsulin-specific T cells in T1D, we isolated human CD4(+) T cell clones to proinsulin from the blood of a donor who had T1D. The clones recognized a naturally processed, HLA DR4-restricted epitope within the first 13 amino acids of the A-chain (A1-13) of human insulin. T cell recognition was dependent on the formation of a vicinal disulfide bond between adjacent cysteine residues at A6 and A7, which did not alter binding of the peptide to HLA DR4. CD4(+) T cell clones that recognized this epitope were isolated from an HLA DR4(+) child with autoantibodies to insulin, and therefore, at risk for T1D, but not from two healthy HLA DR4(+) donors. We define for the first time a novel posttranslational modification that is required for T cell recognition of the insulin A-chain in T1D.
Figures
References
-
- Eisenbarth, G.S. 1986. Type I diabetes mellitus. A chronic autoimmune disease. N. Engl. J. Med. 314:1360–1368. - PubMed
-
- Pugliese, A., and G.S. Eisenbarth. 1996. Human type 1 diabetes mellitus: genetic susceptiblity and resistance. Type 1 Diabetes. Molecular, Cellular, and Clinical Immunology. G.S. Eisenbarth and K.J. Lafferty, editors. Oxford University Press, New York. 134–152.
-
- Narendran, P., S.I. Mannering, and L.C. Harrison. 2003. Proinsulin–a pathogenic autoantigen in type 1 diabetes. Autoimmun. Rev. 2:204–210. - PubMed
-
- Pugliese, A., D. Brown, D. Garza, D. Murchison, M. Zeller, M. Redondo, J. Diez, G.S. Eisenbarth, D.D. Patel, and C. Ricordi. 2001. Self-antigen-presenting cells expressing diabetes-associated autoantigens exist in both thymus and peripheral lymphoid organs. J. Clin. Invest. 107:555–564. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
