

Protection from Alzheimer's-like disease in the mouse by genetic ablation of inducible nitric oxide synthase
- PMID: 16260491
- PMCID: PMC2213235
- DOI: 10.1084/jem.20051529
Protection from Alzheimer's-like disease in the mouse by genetic ablation of inducible nitric oxide synthase
Abstract
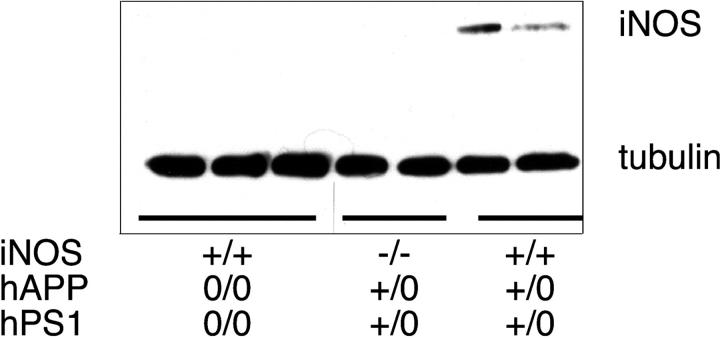
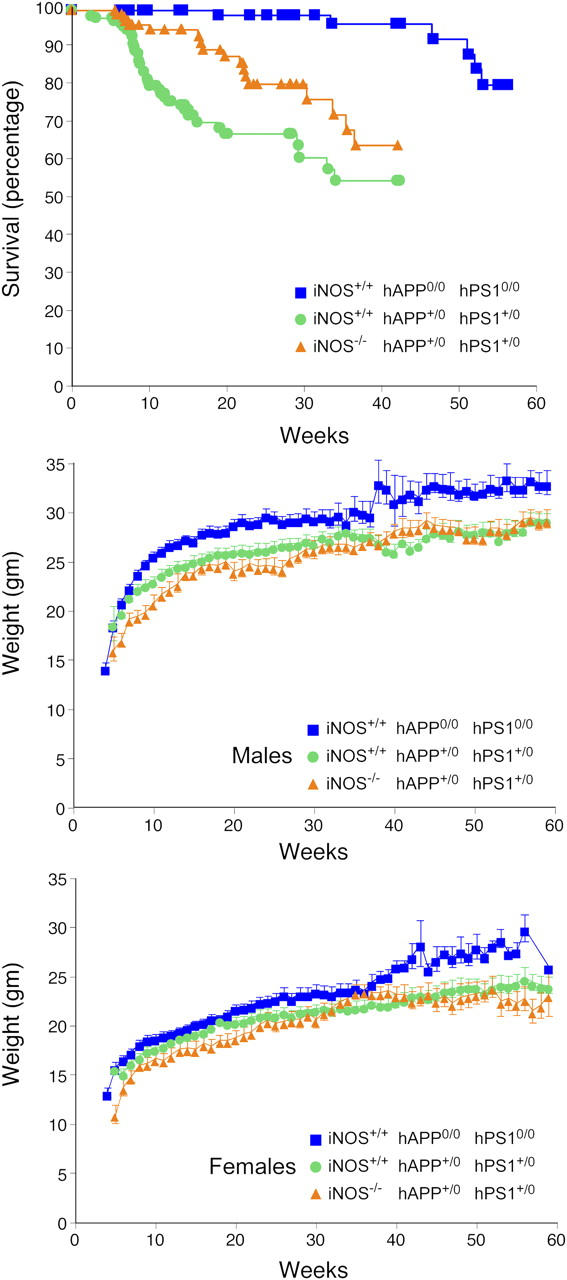
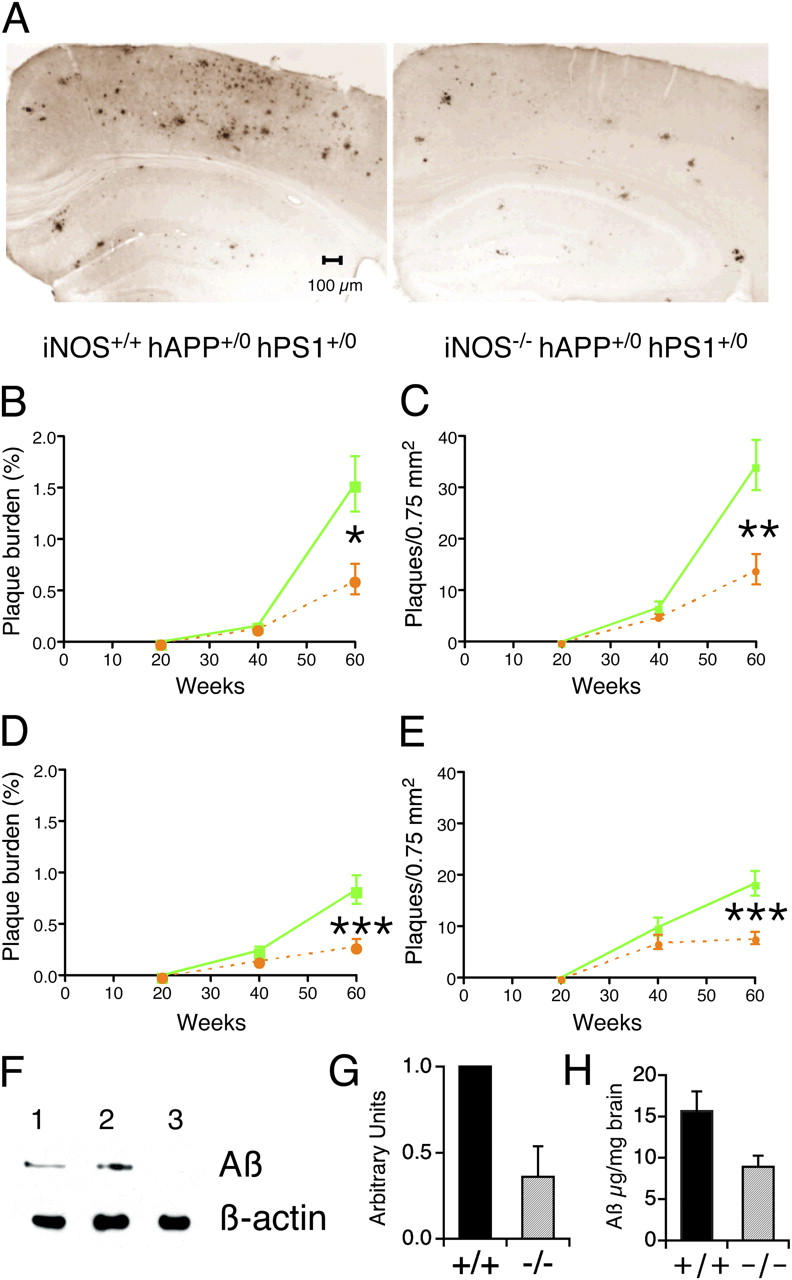
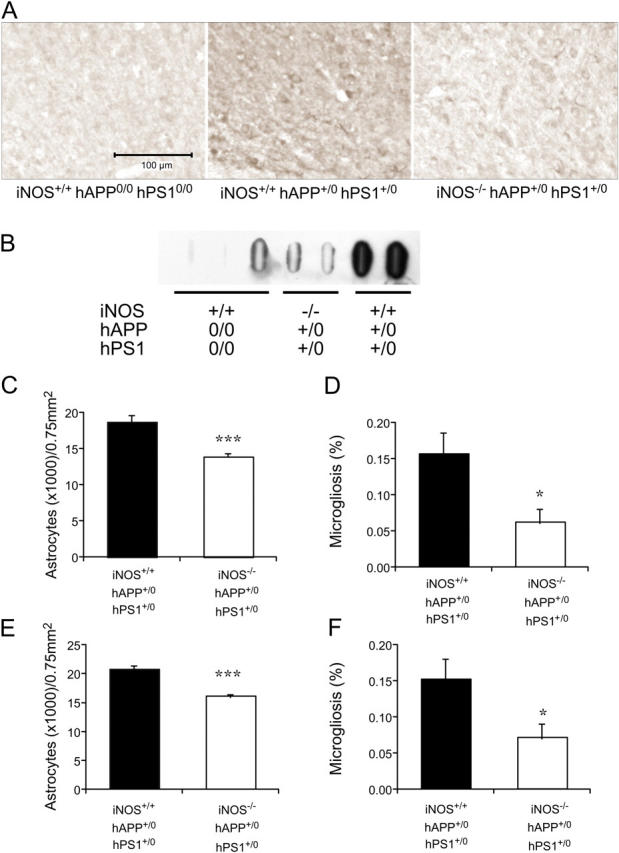
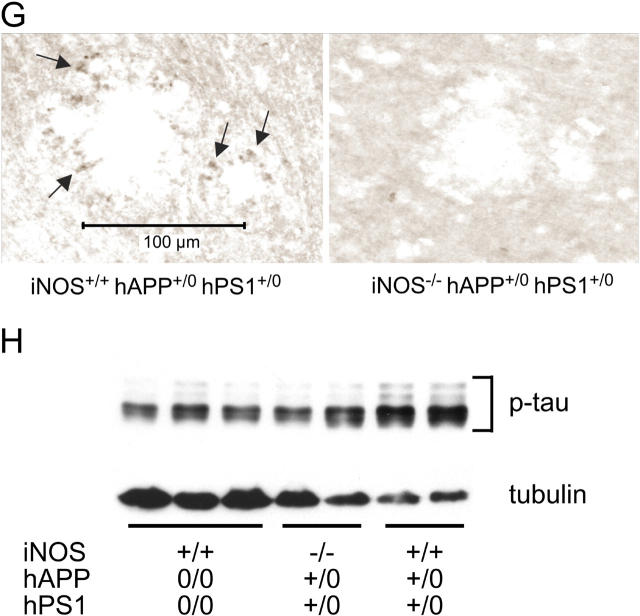
Brains from subjects who have Alzheimer's disease (AD) express inducible nitric oxide synthase (iNOS). We tested the hypothesis that iNOS contributes to AD pathogenesis. Immunoreactive iNOS was detected in brains of mice with AD-like disease resulting from transgenic expression of mutant human beta-amyloid precursor protein (hAPP) and presenilin-1 (hPS1). We bred hAPP-, hPS1-double transgenic mice to be iNOS(+/+) or iNOS(-/-), and compared them with a congenic WT strain. Deficiency of iNOS substantially protected the AD-like mice from premature mortality, cerebral plaque formation, increased beta-amyloid levels, protein tyrosine nitration, astrocytosis, and microgliosis. Thus, iNOS seems to be a major instigator of beta-amyloid deposition and disease progression. Inhibition of iNOS may be a therapeutic option in AD.
Figures
References
-
- Rossi, F., and E. Bianchini. 1996. Synergistic induction of nitric oxide by beta-amyloid and cytokines in astrocytes. Biochem. Biophys. Res. Commun. 225:474–478. - PubMed
-
- Barger, S.W., and A.D. Harmon. 1997. Microglial activation by Alzheimer amyloid precursor protein and modulation by apolipoprotein E. Nature. 388:878–881. - PubMed
-
- Ishii, K., F. Muelhauser, U. Liebl, M. Picard, S. Kuhl, B. Penke, T. Bayer, M. Wiessler, M. Hennerici, K. Beyreuther, et al. 2000. Subacute NO generation induced by Alzheimer's beta-amyloid in the living brain: reversal by inhibition of the inducible NO synthase. FASEB J. 14:1485–1489. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
