

Coupled K+-water flux through the HERG potassium channel measured by an osmotic pulse method
- PMID: 16260841
- PMCID: PMC2266609
- DOI: 10.1085/jgp.200509377
Coupled K+-water flux through the HERG potassium channel measured by an osmotic pulse method
Abstract
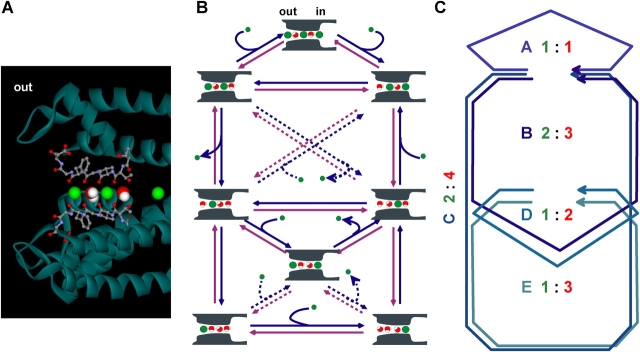
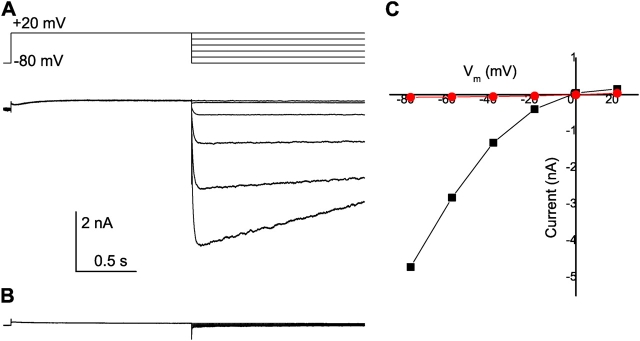
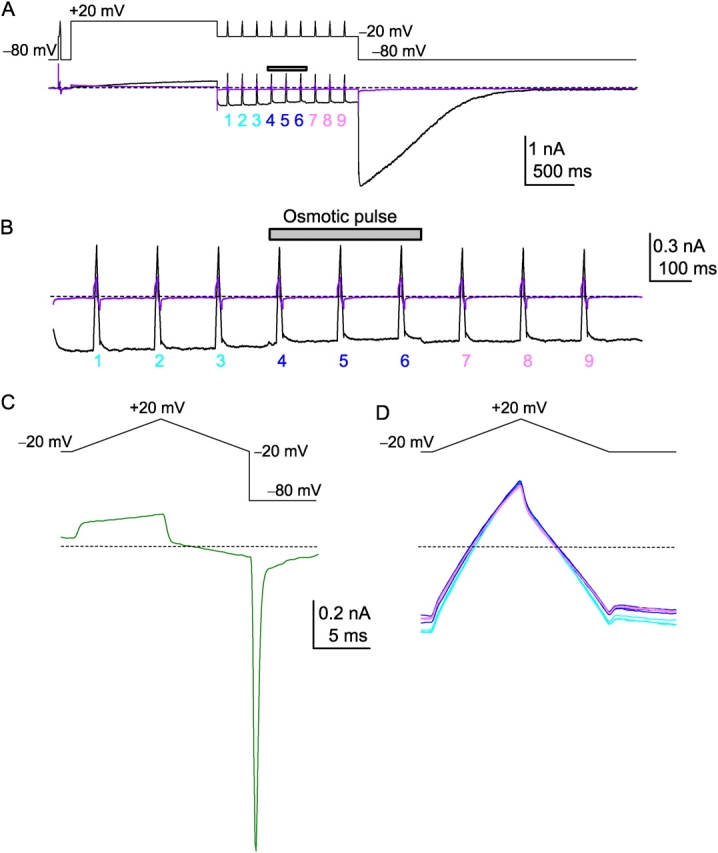
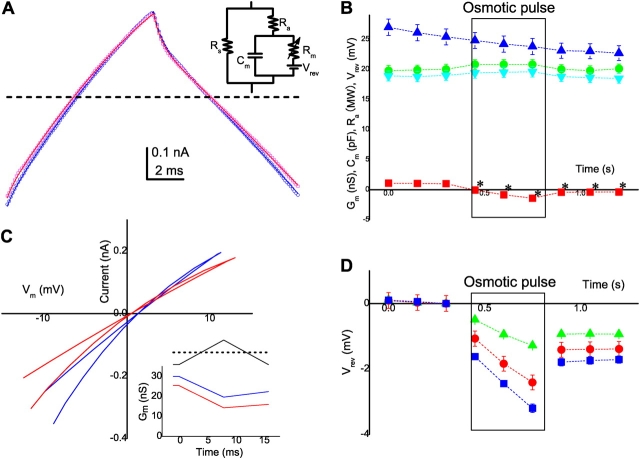
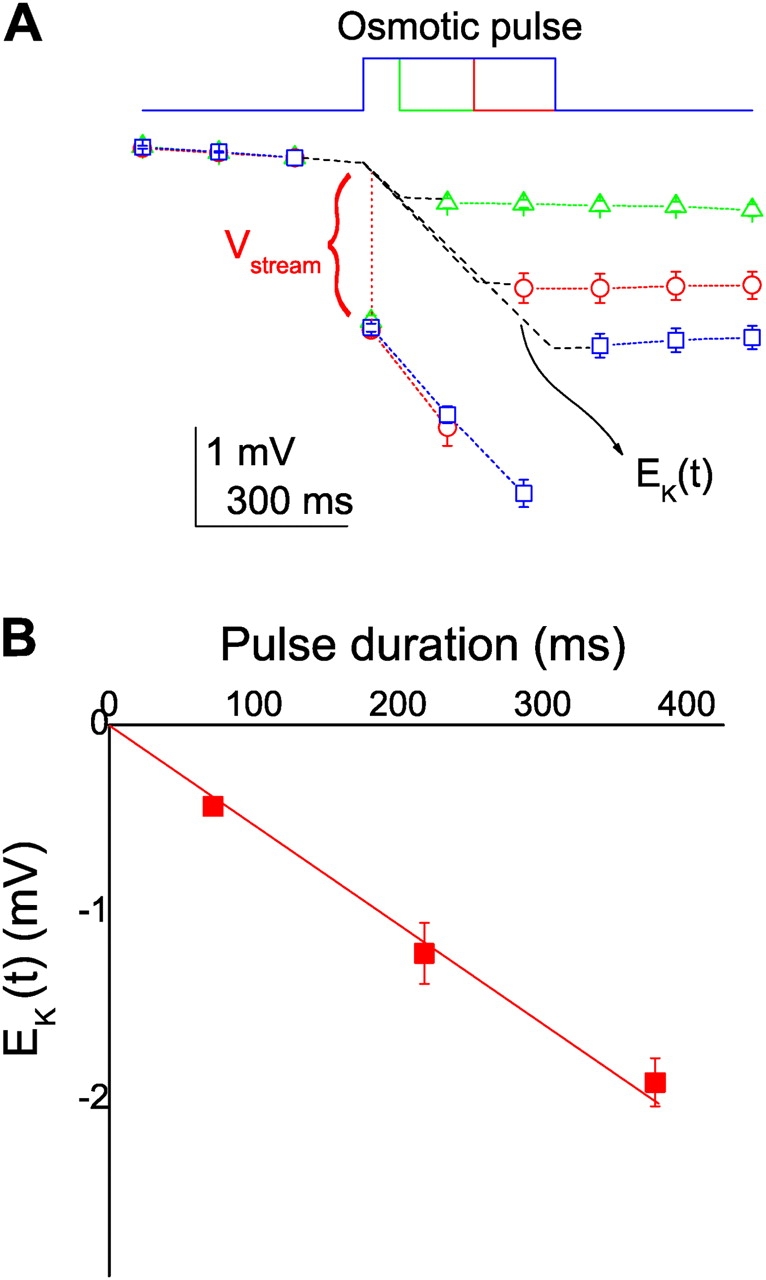
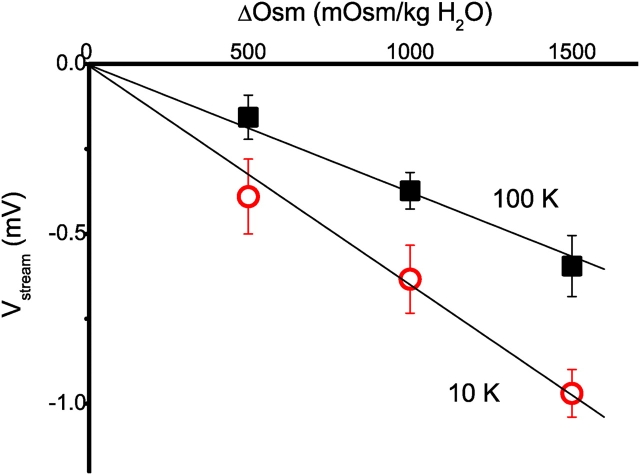
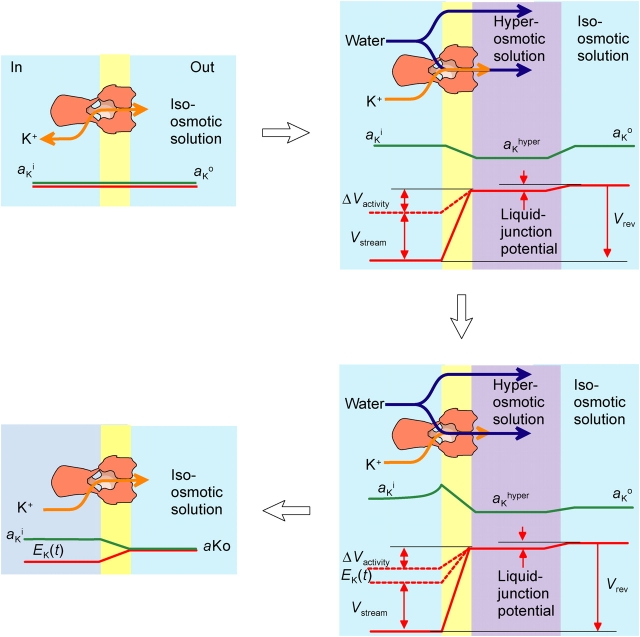
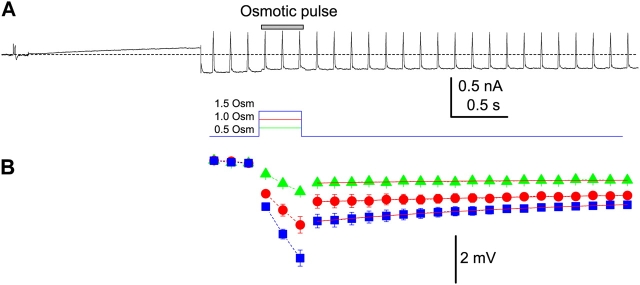
The streaming potential (V(stream)) is a signature feature of ion channels in which permeating ions and water molecules move in a single file. V(stream) provides a quantitative measure of the ion and water flux (the water-ion coupling ratio), the knowledge of which is a prerequisite for elucidating the mechanisms of ion permeation. We have developed a method to measure V(stream) with the whole-cell patch-clamp configuration. A HEK293 cell stably expressing the HERG potassium channel was voltage clamped and exposed to hyperosmotic solutions for short periods of time (<1 s) by an ultrafast solution switching system (the osmotic pulse [quick jump-and-away] method). The reversal potentials were monitored by a series of voltage ramps before, during, and after the osmotic pulse. The shifts of the reversal potentials immediately after the osmotic jump gave V(stream). In symmetrical K+ solutions (10 mM), the V(stream)s measured at different osmolalities showed a linear relationship with a slope of -0.7 mV/DeltaOsm, from which the water-ion coupling ratio (n, the ratio of the flux of water to the flux of cations; Levitt, D.G., S.R. Elias, and J.M. Hautman. 1978. Biochim. Biophys. Acta. 512:436-451) was calculated to be 1.4. In symmetrical 100 mM K+ solutions, the coupling ratio was decreased significantly (n = 0.9), indicating that the permeation process through states with increased ion occupancy became significant. We presented a diagrammatic representation linking the water-ion coupling ratio to the mode of ion permeation and suggested that the coupling ratio of one may represent the least hydrated ion flux in the single-file pore.
Figures
References
-
- Berneche, S., and B. Roux. 2001. Energetics of ion conduction through the K+ channel. Nature. 414:73–77. - PubMed
-
- Finkelstein, A., and O.S. Andersen. 1981. The gramicidin A channel: a review of its permeability characteristics with special reference to the single-file aspect of transport. J. Membr. Biol. 59:155–171. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
