

"Maturational" globin switching in primary primitive erythroid cells
- PMID: 16263786
- PMCID: PMC1895399
- DOI: 10.1182/blood-2005-08-3097
"Maturational" globin switching in primary primitive erythroid cells
Abstract
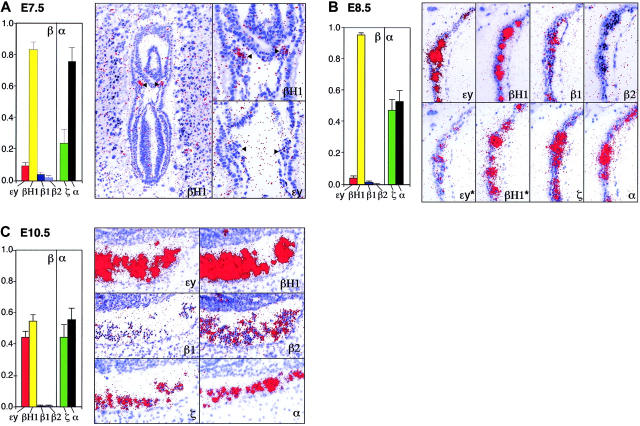
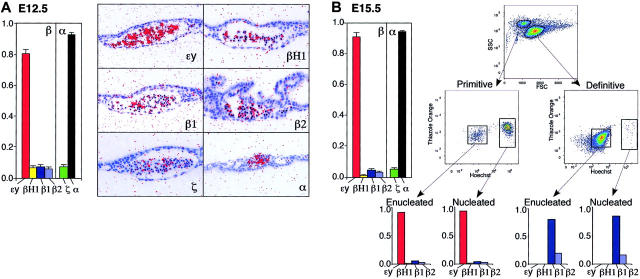
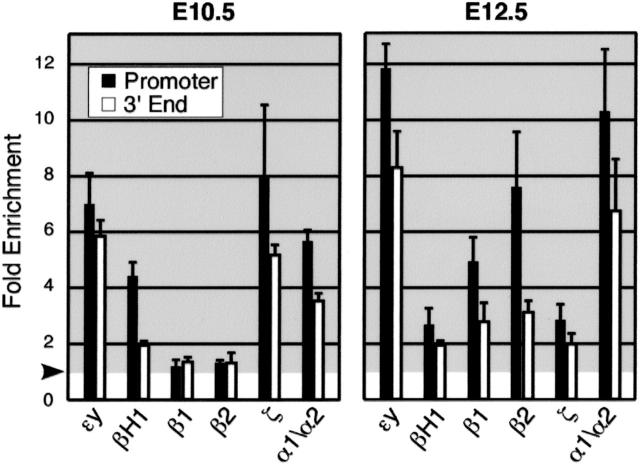
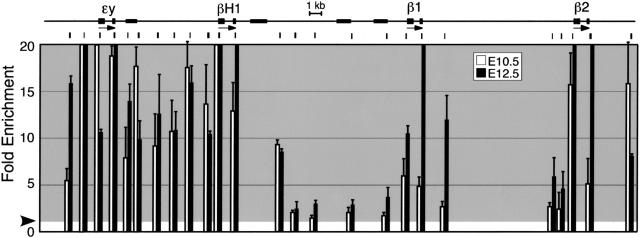
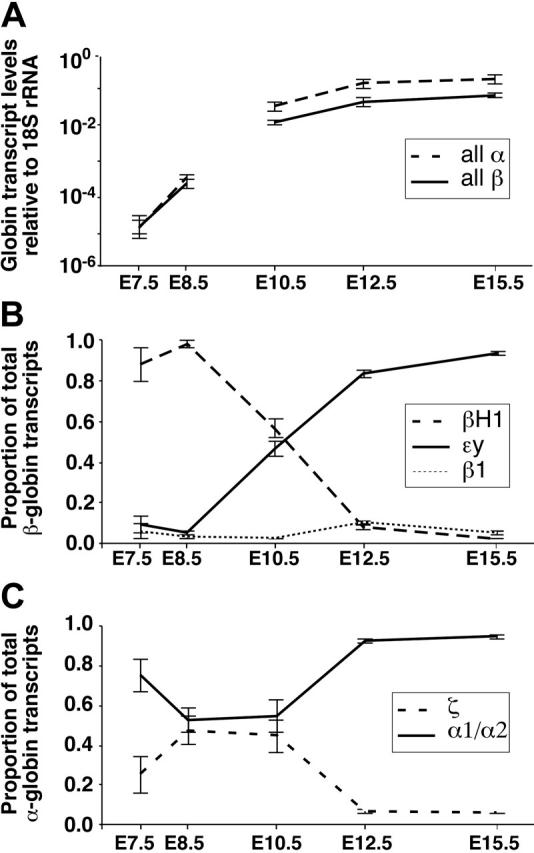
Mammals have 2 distinct erythroid lineages. The primitive erythroid lineage originates in the yolk sac and generates a cohort of large erythroblasts that terminally differentiate in the bloodstream. The definitive erythroid lineage generates smaller enucleated erythrocytes that become the predominant cell in fetal and postnatal circulation. These lineages also have distinct globin expression patterns. Our studies in primary murine primitive erythroid cells indicate that betaH1 is the predominant beta-globin transcript in the early yolk sac. Thus, unlike the human, murine beta-globin genes are not up-regulated in the order of their chromosomal arrangement. As primitive erythroblasts mature from proerythroblasts to reticulocytes, they undergo a betaH1- to epsilony-globin switch, up-regulate adult beta1- and beta2-globins, and down-regulate zeta-globin. These changes in transcript levels correlate with changes in RNA polymerase II density at their promoters and transcribed regions. Furthermore, the epsilony- and betaH1-globin genes in primitive erythroblasts reside within a single large hyperacetylated domain. These data suggest that this "maturational" betaH1- to epsilony-globin switch is dynamically regulated at the transcriptional level. Globin switching during ontogeny is due not only to the sequential appearance of primitive and definitive lineages but also to changes in globin expression as primitive erythroblasts mature in the bloodstream.
Figures
References
-
- Wong PMC, Chung S-H, Reicheld SM, Chui DHK. Hemoglobin switching during murine embryonic development: evidence for two populations of embryonic erythropoietic progenitor cells. Blood. 1986;67: 716-721. - PubMed
-
- Palis J, Robertson S, Kennedy M, Wall C, Keller G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development. 1999;126: 5073-5084. - PubMed
-
- McGrath KE, Koniski AD, Malik J, Palis J. Circulation is established in a stepwise pattern in the mammalian embryo. Blood. 2003;101: 1669-1676. - PubMed
-
- Steiner R, Vogel H. On the kinetics of erythroid cell differentiation in fetal mice, I: microspectrophotometric determination of the hemoglobin content in erythroid cells during gestation. J Cell Physiol. 1973;81: 323-338. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
