

Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum
- PMID: 16263921
- PMCID: PMC1283850
- DOI: 10.1073/pnas.0508467102
Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum
Abstract
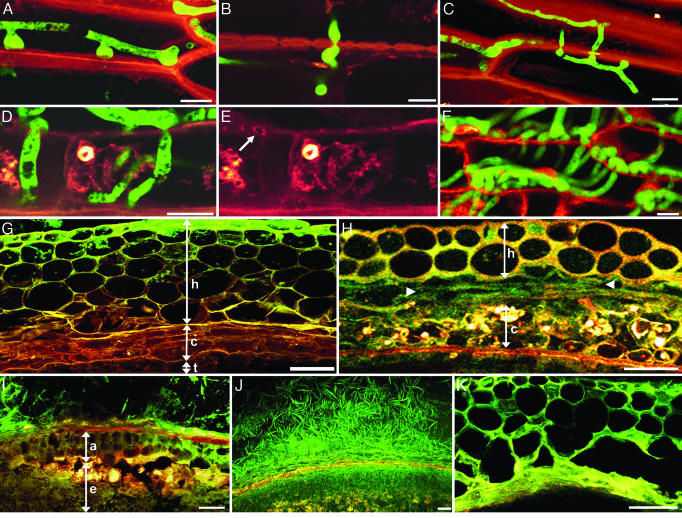
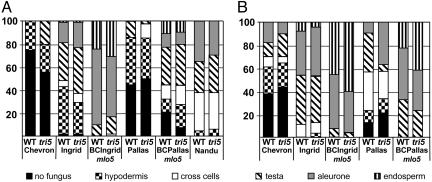
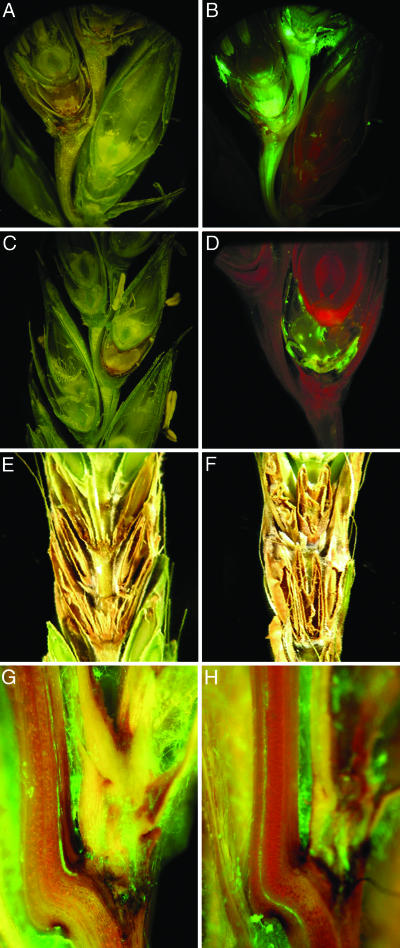
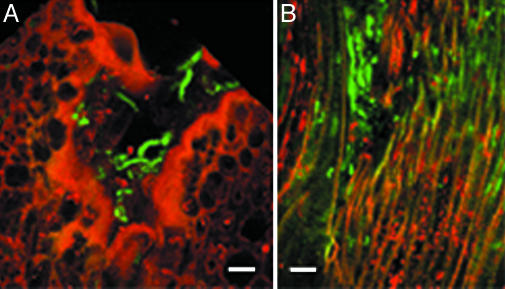
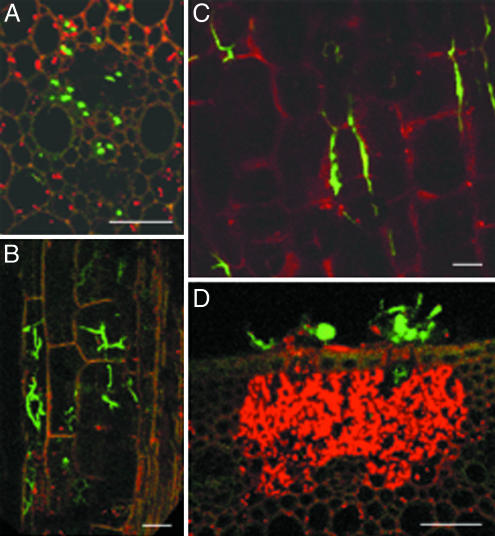
Fusarium head blight epidemics of wheat and barley cause heavy economic losses to farmers due to yield decreases and production of mycotoxin that renders the grain useless for flour and malt products. No highly resistant cultivars are available at present. Hyphae of germinating fungal spores use different paths of infection: After germination at the extruded tip of an ovary, the hyphae travel along the epicarp in the space between the lemma and palea. Infection of the developing kernel proceeds through the epicarp, successively destroying the layers of the fruit coat and finally the starch and protein accumulating endosperm. Hyphae reaching the rachis proceed to apically located developing kernels. Using a constitutively green fluorescence protein-expressing Fusarium wild-type strain, and its knockout mutant, preventing trichothecene synthesis, we demonstrate that trichothecenes are not a virulence factor during infection through the fruit coat. In the absence of trichothecenes, the fungus is blocked by the development of heavy cell wall thickenings in the rachis node of Nandu wheat, a defense inhibited by the mycotoxin. In barley hyphae of both wild-type and the trichothecene knockout mutant, are inhibited at the rachis node and rachilla, limiting infection of adjacent florets through the phloem and along the surface of the rachis. Effective resistance to Fusarium head blight requires expression of genes that combat these different pathways of infection.
Figures
Similar articles
-
Developing kernel and rachis node induce the trichothecene pathway of Fusarium graminearum during wheat head infection.Mol Plant Microbe Interact. 2009 Aug;22(8):899-908. doi: 10.1094/MPMI-22-8-0899. Mol Plant Microbe Interact. 2009. PMID: 19589066
-
Transcriptome analysis of trichothecene-induced gene expression in barley.Mol Plant Microbe Interact. 2007 Nov;20(11):1364-75. doi: 10.1094/MPMI-20-11-1364. Mol Plant Microbe Interact. 2007. PMID: 17977148
-
Metabolic profiling of wheat rachis node infection by Fusarium graminearum - decoding deoxynivalenol-dependent susceptibility.New Phytol. 2019 Jan;221(1):459-469. doi: 10.1111/nph.15377. Epub 2018 Aug 7. New Phytol. 2019. PMID: 30084118
-
Management and resistance in wheat and barley to fusarium head blight.Annu Rev Phytopathol. 2004;42:135-61. doi: 10.1146/annurev.phyto.42.040803.140340. Annu Rev Phytopathol. 2004. PMID: 15283663 Review.
-
Resistance in wheat to Fusarium infection and trichothecene formation.Toxicol Lett. 2004 Oct 10;153(1):37-46. doi: 10.1016/j.toxlet.2004.04.044. Toxicol Lett. 2004. PMID: 15342079 Review.
Cited by
-
Discovery of a susceptibility factor for Fusarium head blight on chromosome 7A of wheat.Theor Appl Genet. 2021 Jul;134(7):2273-2289. doi: 10.1007/s00122-021-03825-y. Epub 2021 Apr 8. Theor Appl Genet. 2021. PMID: 33834252
-
Transcriptome-based discovery of pathways and genes related to resistance against Fusarium head blight in wheat landrace Wangshuibai.BMC Genomics. 2013 Mar 21;14:197. doi: 10.1186/1471-2164-14-197. BMC Genomics. 2013. PMID: 23514540 Free PMC article.
-
Regulation of TRI5 expression and deoxynivalenol biosynthesis by a long non-coding RNA in Fusarium graminearum.Nat Commun. 2024 Feb 9;15(1):1216. doi: 10.1038/s41467-024-45502-w. Nat Commun. 2024. PMID: 38332031 Free PMC article.
-
Changes in Antioxidant System during Grain Development of Wheat (Triticum aestivum L.) and Relationship with Protein Composition under FHB Stress.Pathogens. 2019 Dec 23;9(1):17. doi: 10.3390/pathogens9010017. Pathogens. 2019. PMID: 31878029 Free PMC article.
-
Integrated transcriptome and hormone profiling highlight the role of multiple phytohormone pathways in wheat resistance against fusarium head blight.PLoS One. 2018 Nov 7;13(11):e0207036. doi: 10.1371/journal.pone.0207036. eCollection 2018. PLoS One. 2018. PMID: 30403737 Free PMC article.
References
-
- Windels, C. E. (2000) Phytopathology 90, 17-21. - PubMed
-
- National Agricultural Statistics Service (1993-1999) Annual Reports: Minnesota Agricultural Statistics, Minnesota Office, St. Paul and North Dakota Agricultural Statistics, North Dakota Office, Fargo (United States Department of Agriculture, Washington, DC).
-
- Placinta, C. M., D'Mello, J. P. F. & Macdonald, A. M. C. (1999) Anim. Feed Sci. Tech. 78, 21-37.
-
- Bai, G. & Shaner, G. (2004) Annu. Rev. Phytopathol. 42, 135-161. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
