

Characterization of the metabolic shift between oxidative and fermentative growth in Saccharomyces cerevisiae by comparative 13C flux analysis
- PMID: 16269086
- PMCID: PMC1291395
- DOI: 10.1186/1475-2859-4-30
Characterization of the metabolic shift between oxidative and fermentative growth in Saccharomyces cerevisiae by comparative 13C flux analysis
Abstract
Background: One of the most fascinating properties of the biotechnologically important organism Saccharomyces cerevisiae is its ability to perform simultaneous respiration and fermentation at high growth rate even under fully aerobic conditions. In the present work, this Crabtree effect called phenomenon was investigated in detail by comparative 13C metabolic flux analysis of S. cerevisiae growing under purely oxidative, respiro-fermentative and predominantly fermentative conditions.
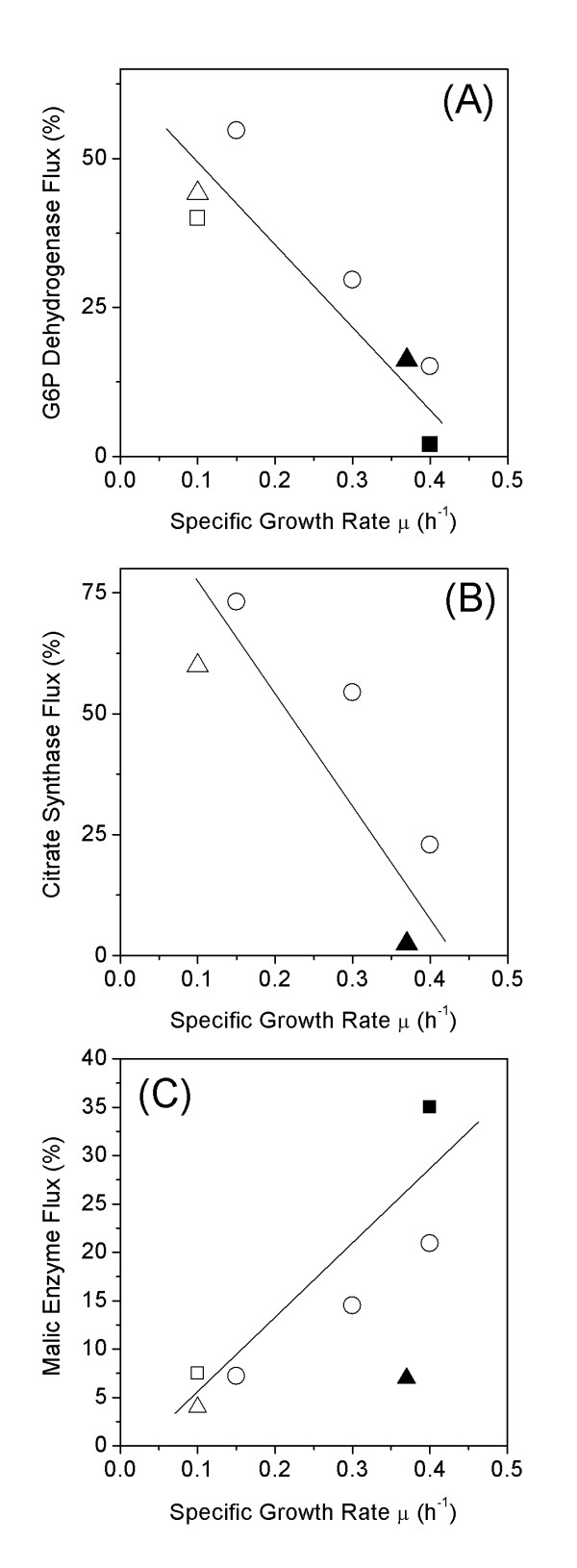
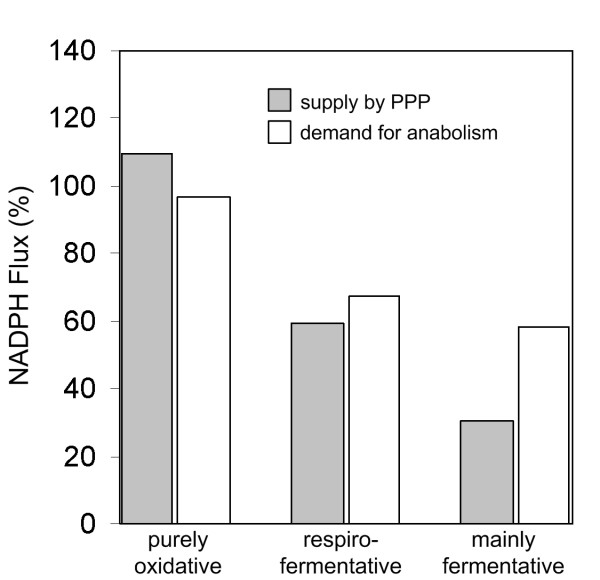
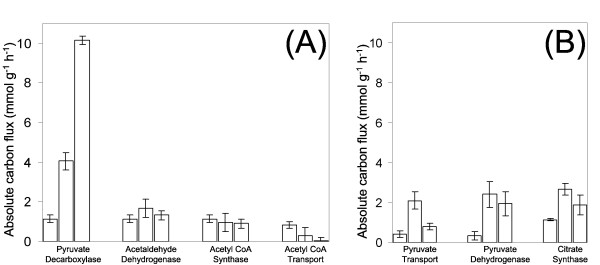
Results: The metabolic shift from oxidative to fermentative growth was accompanied by complex changes of carbon flux throughout the whole central metabolism. This involved a flux redirection from the pentose phosphate pathway (PPP) towards glycolysis, an increased flux through pyruvate carboxylase, the fermentative pathways and malic enzyme, a flux decrease through the TCA cycle, and a partial relocation of alanine biosynthesis from the mitochondrion to the cytosol. S. cerevisiae exhibited a by-pass of pyruvate dehydrogenase in all physiological regimes. During oxidative growth this by-pass was mainly provided via pyruvate decarboxylase, acetaldehyde dehydrogenase, acetyl-CoA synthase and transport of acetyl-CoA into the mitochondrion. During fermentative growth this route, however, was saturated due to limited enzyme capacity. Under these conditions the cells exhibited high carbon flux through a chain of reactions involving pyruvate carboxylase, the oxaloacetate transporter and malic enzyme. During purely oxidative growth the PPP alone was sufficient to completely supply NADPH for anabolism. During fermentation, it provided only 60 % of the required NADPH.
Conclusion: We conclude that, in order to overcome the limited capacity of pyruvate dehydrogenase, S. cerevisiae possesses different metabolic by-passes to channel carbon into the mitochondrion. This involves the conversion of cytosolic pyruvate either into acetyl CoA or oxaloacetate followed by intercompartmental transport of these metabolites. During oxidative growth mainly the NAD specific isoforms of acetaldehyde dehydrogenase and isocitrate dehydrogenase catalyze the corresponding reactions in S. cerevisiae, whereas NADPH supply under fermentative conditions involves significant contribution of sources other than the PPP such as e. g. NADPH specific acetaldehyde dehydrogenase or isocitrate dehydrogenase.
Figures



References
-
- Kato A, Nakamura S, Ibrahim H, Matsumi T, Tsumiyama C, Kato M. Production of genetically modified lysozymes having extreme heat stability and antimicrobial activity against gram negative bacteria in yeast and in plant. Nahrung. 1998;42:128–130. doi: 10.1002/(SICI)1521-3803(199808)42:03/04<128::AID-FOOD128>3.3.CO;2-Q. - DOI - PubMed
-
- Kim MD, Lee TH, Lim HK, Seo JH. Production of antithrombotic hirudin in GAL1-disrupted Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2004;65:259–262. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
