

Culture-dependent and culture-independent diversity within the obligate marine actinomycete genus Salinispora
- PMID: 16269737
- PMCID: PMC1287694
- DOI: 10.1128/AEM.71.11.7019-7028.2005
Culture-dependent and culture-independent diversity within the obligate marine actinomycete genus Salinispora
Abstract
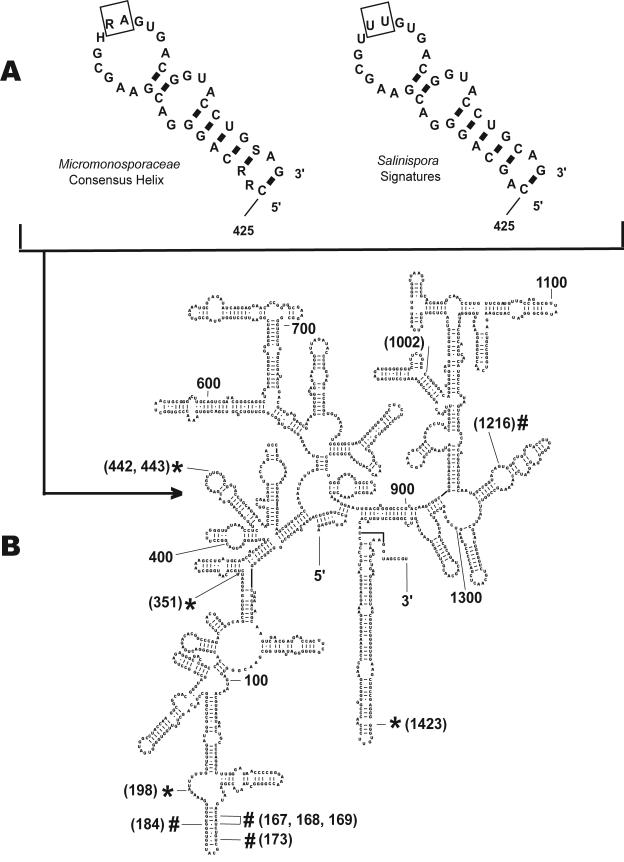
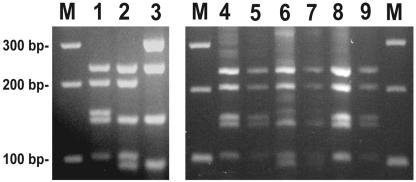
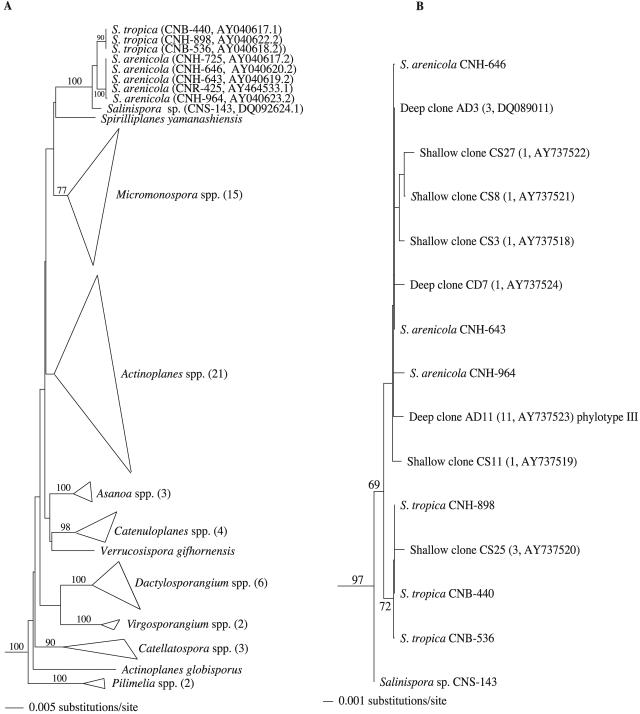
Salinispora is the first obligate marine genus within the order Actinomycetales and a productive source of biologically active secondary metabolites. Despite a worldwide, tropical or subtropical distribution in marine sediments, only two Salinispora species have thus far been cultivated, suggesting limited species-level diversity. To further explore Salinispora diversity and distributions, the phylogenetic diversity of more than 350 strains isolated from sediments collected around the Bahamas was examined, including strains cultured using new enrichment methods. A culture-independent method, using a Salinispora-specific seminested PCR technique, was used to detect Salinispora from environmental DNA and estimate diversity. Overall, the 16S rRNA gene sequence diversity of cultured strains agreed well with that detected in the environmental clone libraries. Despite extensive effort, no new species level diversity was detected, and 97% of the 105 strains examined by restriction fragment length polymorphism belonged to one phylotype (S. arenicola). New intraspecific diversity was detected in the libraries, including an abundant new phylotype that has yet to be cultured, and a new depth record of 1,100 m was established for the genus. PCR-introduced error, primarily from Taq polymerase, significantly increased clone library sequence diversity and, if not masked from the analyses, would have led to an overestimation of total diversity. An environmental DNA extraction method specific for vegetative cells provided evidence for active actinomycete growth in marine sediments while indicating that a majority of sediment samples contained predominantly Salinispora spores at concentrations that could not be detected in environmental clone libraries. Challenges involved with the direct sequence-based detection of spore-forming microorganisms in environmental samples are discussed.
Figures
Similar articles
-
Biogeography of the marine actinomycete Salinispora.Environ Microbiol. 2006 Nov;8(11):1881-8. doi: 10.1111/j.1462-2920.2006.01093.x. Environ Microbiol. 2006. PMID: 17014488
-
Salinispora arenicola from temperate marine sediments: new intra-species variations and atypical distribution of secondary metabolic genes.Antonie Van Leeuwenhoek. 2014 Jan;105(1):207-19. doi: 10.1007/s10482-013-0067-2. Epub 2013 Nov 1. Antonie Van Leeuwenhoek. 2014. PMID: 24179039
-
Culturable marine actinomycete diversity from tropical Pacific Ocean sediments.Environ Microbiol. 2005 Jul;7(7):1039-48. doi: 10.1111/j.1462-2920.2005.00785.x. Environ Microbiol. 2005. PMID: 15946301
-
Bioactive natural products from the genus Salinospora: a review.Arch Pharm Res. 2020 Dec;43(12):1230-1258. doi: 10.1007/s12272-020-01288-1. Epub 2020 Nov 25. Arch Pharm Res. 2020. PMID: 33237436 Review.
-
Physiology of some actinomycete genera.Res Microbiol. 1993 Oct;144(8):657-60. doi: 10.1016/0923-2508(93)90070-i. Res Microbiol. 1993. PMID: 8140284 Review.
Cited by
-
Antarctic sponges from the Terra Nova Bay (Ross Sea) host a diversified bacterial community.Sci Rep. 2019 Nov 6;9(1):16135. doi: 10.1038/s41598-019-52491-0. Sci Rep. 2019. PMID: 31695084 Free PMC article.
-
An assessment of actinobacterial diversity in the marine environment.Antonie Van Leeuwenhoek. 2008 Jun;94(1):51-62. doi: 10.1007/s10482-008-9239-x. Epub 2008 May 24. Antonie Van Leeuwenhoek. 2008. PMID: 18500568 Free PMC article.
-
Bioprospecting marine actinomycetes for antileishmanial drugs: current perspectives and future prospects.Heliyon. 2021 Aug 2;7(8):e07710. doi: 10.1016/j.heliyon.2021.e07710. eCollection 2021 Aug. Heliyon. 2021. PMID: 34409179 Free PMC article. Review.
-
Bioactive compounds from marine actinomycetes.Indian J Microbiol. 2008 Dec;48(4):410-31. doi: 10.1007/s12088-008-0052-z. Epub 2009 Jan 8. Indian J Microbiol. 2008. PMID: 23100742 Free PMC article.
-
Exploring the diversity and metabolic potential of actinomycetes from temperate marine sediments from Newfoundland, Canada.J Ind Microbiol Biotechnol. 2015 Jan;42(1):57-72. doi: 10.1007/s10295-014-1529-x. Epub 2014 Nov 5. J Ind Microbiol Biotechnol. 2015. PMID: 25371290
References
-
- Acinas, S. G., V. Klepac-Ceraj, D. E. Hunt, C. Pharino, I. C. Ceraj, D. L. Distel, and M. F. Poltz. 2004. Fine-scale phylogenetic architecture of a complex bacterial community. Nature 430:551-554. - PubMed
-
- Berdy, J. 2005. Bioactive microbial metabolites: a personal view. J. Antibiot. 58:1-26. - PubMed
-
- Bracho, M. A., A. Moya, and E. Barrio. 1998. Contribution of Taq polymerase-induced errors to the estimation of RNA virus diversity. J. Gen. Virol. 79:2921-2928. - PubMed
-
- Cannone, J. J., S. Subramanian, M. N. Schnare, J. R. Collett, L. M. D'Souza, Y. Du, B. Feng, N. Lin, L. V. Madabusi, K. M. Muller, N. Pande, Z. Shang, N. Yu, and R. R. Gutell. 2002. The Comparative RNA Web (CRW) Site: an online database of comparative sequence and structure information for ribosomal, intron, and other RNAs. BioMed. Central Bioinform. 3. [Online.] - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
