

Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron
- PMID: 16270029
- PMCID: PMC1356305
- DOI: 10.1038/sj.emboj.7600864
Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron
Abstract
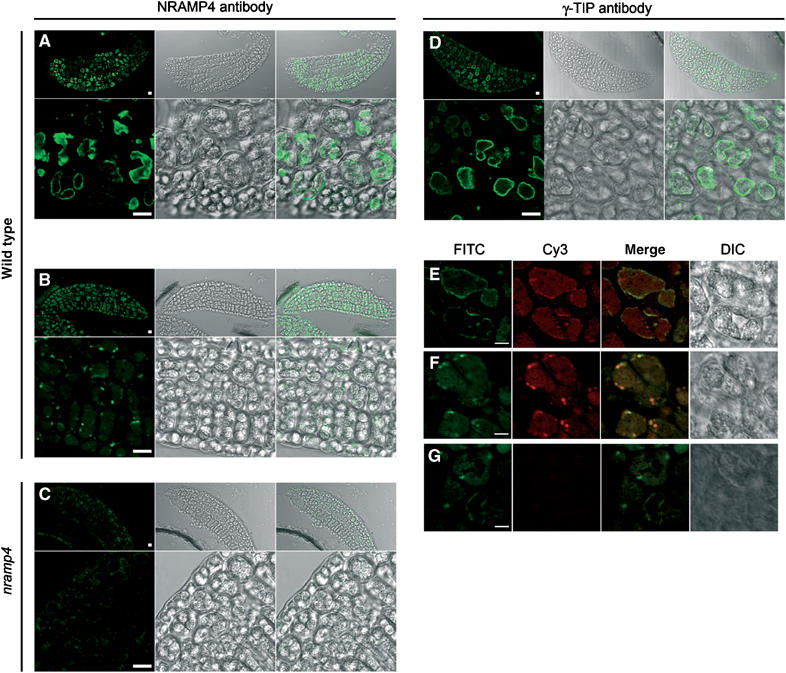
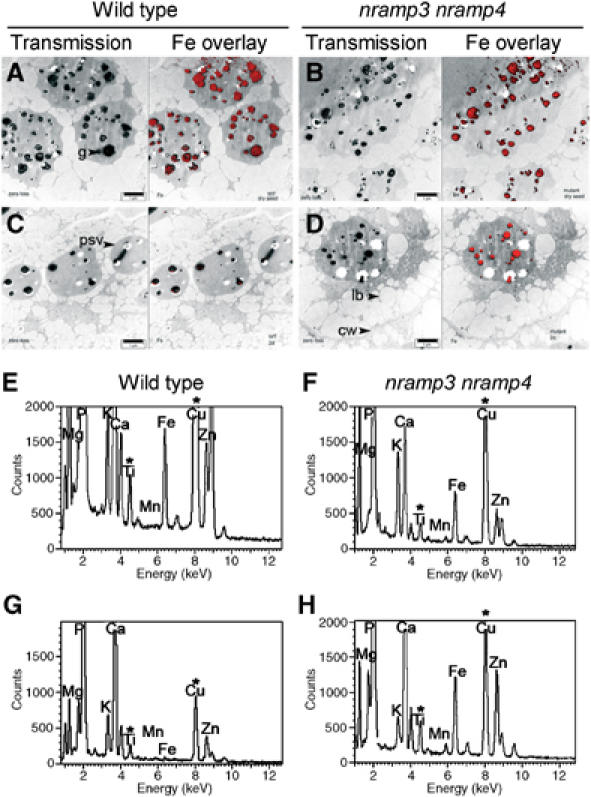
Iron (Fe) is necessary for all living cells, but its bioavailability is often limited. Fe deficiency limits agriculture in many areas and affects over a billion human beings worldwide. In mammals, NRAMP2/DMT1/DCT1 was identified as a major pathway for Fe acquisition and recycling. In plants, AtNRAMP3 and AtNRAMP4 are induced under Fe deficiency. The similitude of AtNRAMP3 and AtNRAMP4 expression patterns and their common targeting to the vacuole, together with the lack of obvious phenotype in nramp3-1 and nramp4-1 single knockout mutants, suggested a functional redundancy. Indeed, the germination of nramp3 nramp4 double mutants is arrested under low Fe nutrition and fully rescued by high Fe supply. Mutant seeds have wild type Fe content, but fail to retrieve Fe from the vacuolar globoids. Our work thus identifies for the first time the vacuole as an essential compartment for Fe storage in seeds. Our data indicate that mobilization of vacuolar Fe stores by AtNRAMP3 and AtNRAMP4 is crucial to support Arabidopsis early development until efficient systems for Fe acquisition from the soil take over.
Figures
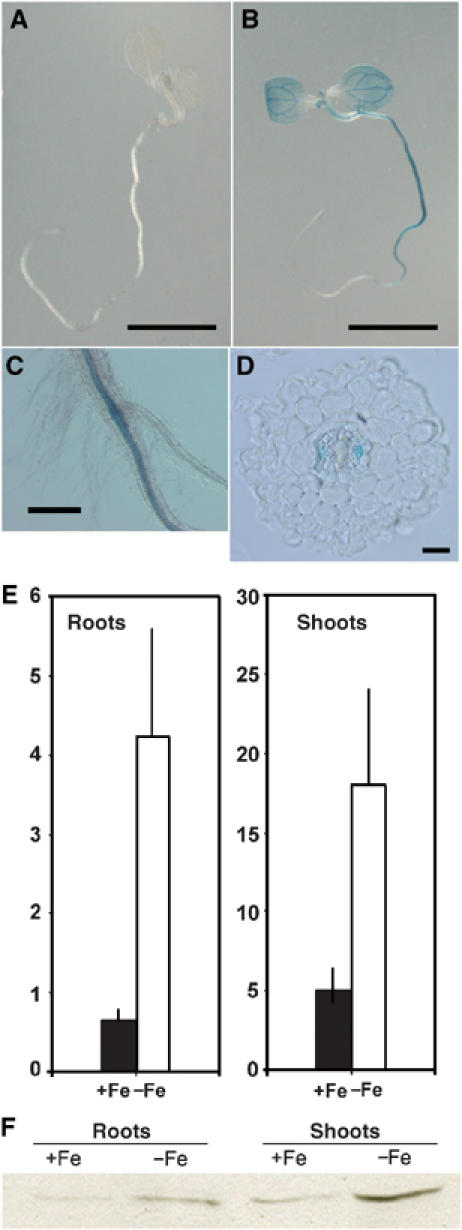
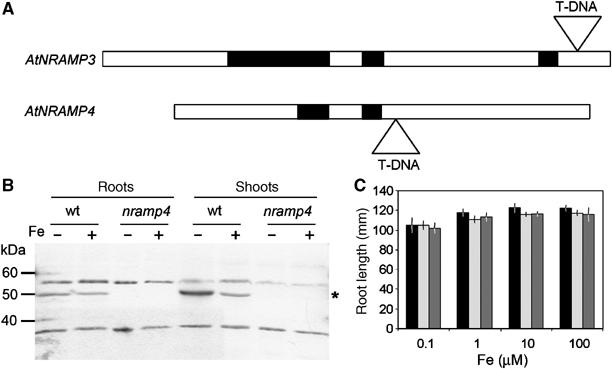
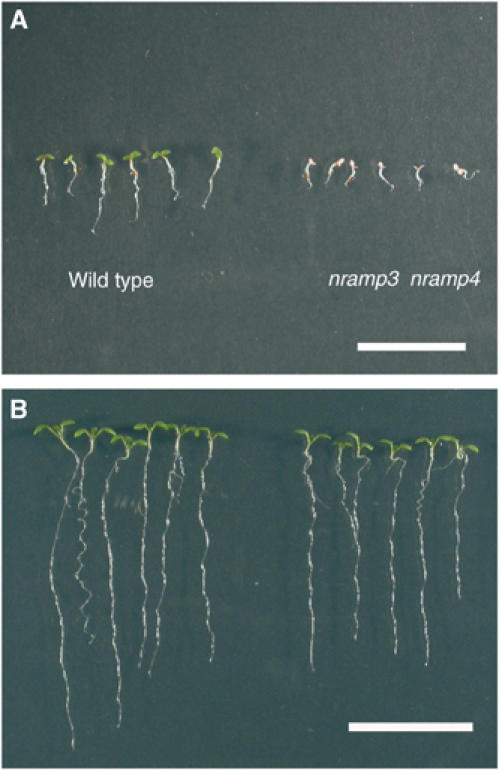
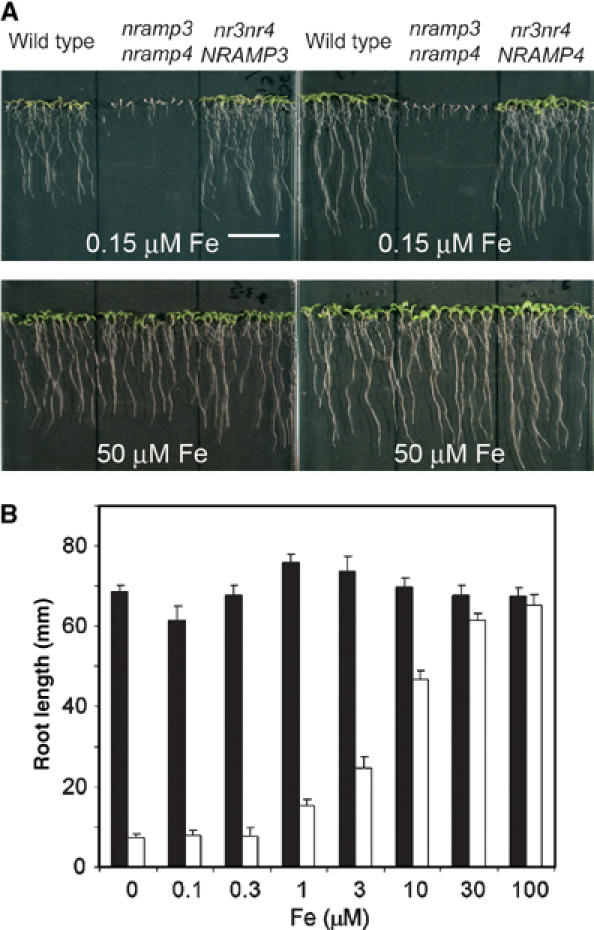


References
-
- Beeckman T, Viane R (1999) Embedding thin plant specimens for oriented sectioning. Biotechn Histochem 75: 23–26 - PubMed
-
- Bensmihen S, To A, Lambert G, Kroj T, Giraudat J, Parcy F (2004) Analysis of an activated ABI5 allele using a new selection method for transgenic Arabidopsisseeds. FEBS Lett 561: 127–131 - PubMed
-
- Bereczky Z, Wang HY, Schubert V, Ganal M, Bauer P (2003) Differential regulation of nramp and irt metal transporter genes in wild type and iron uptake mutants of tomato. J Biol Chem 278: 24697–24704 - PubMed
-
- Bolte S, Brown S, Satiat-Jeunemaitre B (2004) The N-myristoylated Rab-GTPase m-Rabmc is involved in post-Golgi trafficking events to the lytic vacuole in plant cells. J Cell Sci 117: 943–954 - PubMed
-
- Buttrose MS (1978) Manganese and iron in globoid crystals of protein bodies from Avena and Casuarina. Aust J Plant Physiol 5: 631–639
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
