

Review
doi: 10.1523/JNEUROSCI.4119-05.2005.
Epigenetic mechanisms and gene networks in the nervous system
Affiliations
- PMID: 16280577
- PMCID: PMC6725821
- DOI: 10.1523/JNEUROSCI.4119-05.2005
Item in Clipboard
Review
Epigenetic mechanisms and gene networks in the nervous system
J Neurosci.
.
No abstract available
Figures
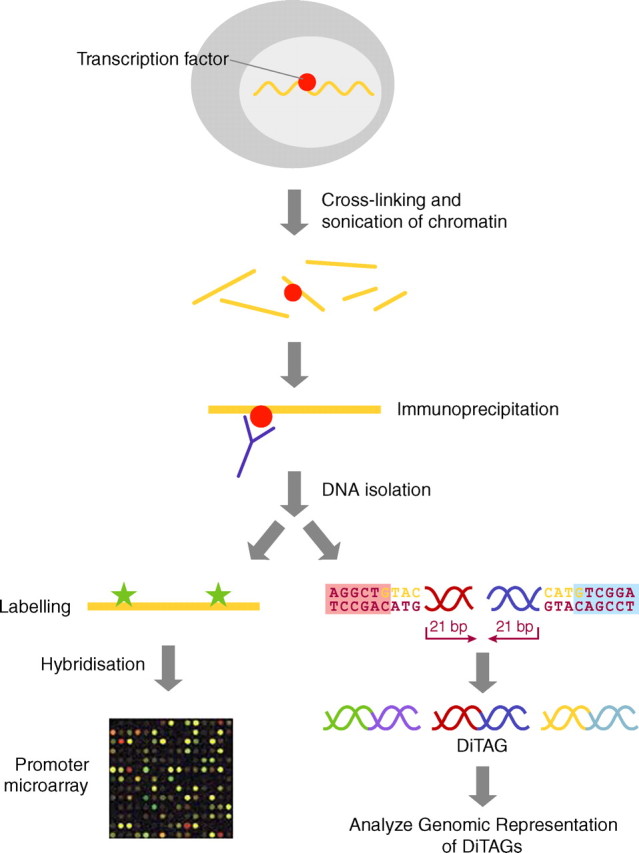
Schematic of hybridization (ChIP-on-chip) and sequence (SACO)-based genomic screens for transcription factor binding sites. For the details of the methods, see Odom et al. (2004) and Impey et al. (2004). Figure adapted from Servitja and Ferrer (2004)
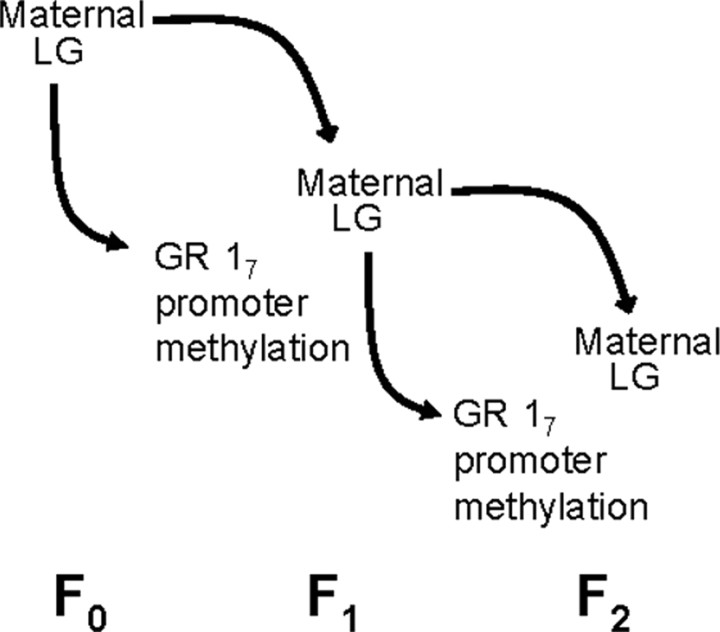
Illustration of the behavioral transmission of epigenetic modifications across generations via maternal care. Variations in maternal LG are transmitted from mother (F0) to female offspring (F1), and differential methylation of the 17 promoter region of the GR is likewise induced by F0 LG. Variation in LG of female offspring (F1) is then transmitted to the F2 generation, as are patterns of GR methylation.
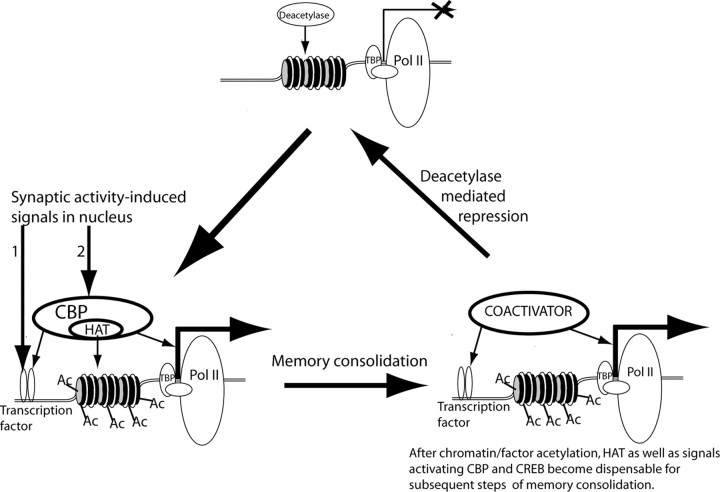
Acetylation is a critical requirement for long-term memory consolidation. The suggested model takes into account five observations: (1) NMDA receptor-dependent neuronal activation is required for memory formation (Davis et al., 1992), (2) neuronal activity-dependent transcription is required for memory consolidation (Andrew, 1980; Davis and Squire, 1984), (3) NMDA-dependent transcription requires the concurrent activation of transcription factors such as CREB and coactivators such as CBP (Chawla et al., 1998; Impey et al., 2002), (4) behaviorally induced CREB phosphorylation is transient and does not correlate with peak gene induction (Stanciu et al., 2001), and (5) CBP acetyltransferase activity is required for long-term memory consolidation but not for short-term memory (Korzus et al., 2004). This model suggests that CBP-mediated histone acetylation is a limiting step in the transcriptional activation controlling memory consolidation. Initial steps include induction of transient CREB phosphorylation, CBP activation, and CBP-mediated histone acetylation at a specific transcriptional unit in response to the initial learning event. Subsequently, the prolonged elevated transcription required for memory consolidation could be maintained by CBP and CREB phosphorylation-independent nuclear mechanisms, even after signals to CREB and CBP are no longer present. This ongoing transcription would remain open until the competing deacetylase-dependent mechanisms suppress transcription (modified from Korzus et al., 2004). Ac, Acetyl groups; Pol II, RNA polymerase II; TBP, TATA-binding protein.
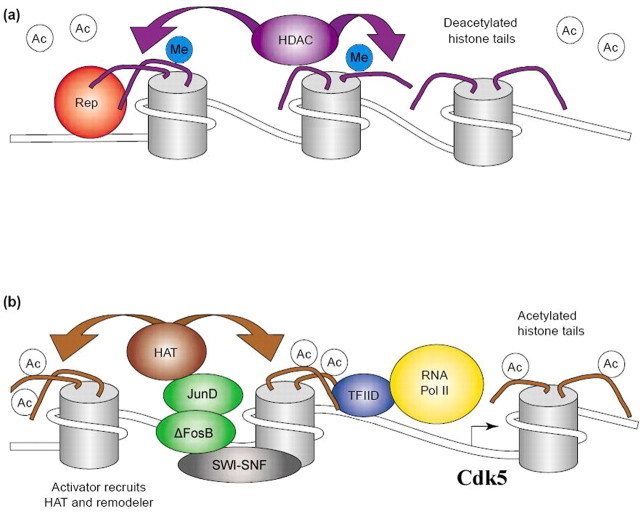
Scheme of proposed chromatin remodeling events at a cocaine-activated gene. A shows the repressed state of chromatin, in which a site-specific repressor (Rep) recruits an HDAC complex, which removes acetyl groups (Ac) from histone N-terminal tails. Gene inactivation may also involve other modifications, such as methylation (Me) of histone tails. B shows the active state of chromatin around a cocaine-activated gene (e.g., Cdk5), in which a cocaine-induced transcriptional activator (e.g., ΔFosB-JunD) recruits a HAT and chromatin remodeling complex (SWI-SNF), which induces acetylation (and perhaps demethylation) of histone tails and repositioning of nucleosomes. These actions facilitate the binding of general transcription factors and the basal transcriptional apparatus [e.g., transcription factor IID (TFIID) and RNA polymerase II (Pol II)] to the promoter.
References
-
- Abel T, Kandel E (1998) Positive and negative regulatory mechanisms that mediate long-term memory storage. Brain Res Brain Res Rev 26: 360-378. - PubMed
-
- Alarcon JM, Malleret G, Touzani K, Vronskaya S, Ishii S, Kandel ER, Barco A (2004) Chromatin acetylation, memory, and LTP are impaired in CBP+/- mice: a model for the cognitive deficit in Rubinstein-Taybi syndrome and its amelioration. Neuron 42: 947-959. - PubMed
-
- Andrew RJ (1980) The functional organization of phases of memory consolidation. In: Advances in the study of behaviour (Hinde RA, Beer C, Bunsel M, eds), pp 337-367. New York: Academic.
-
- Bear MF, Malenka RC (1994) Synaptic plasticity: LTP and LTD. Curr Opin Neurobiol 4: 389-399. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources