

Interactions with pocket proteins contribute to the role of human papillomavirus type 16 E7 in the papillomavirus life cycle
- PMID: 16282477
- PMCID: PMC1287546
- DOI: 10.1128/JVI.79.23.14769-14780.2005
Interactions with pocket proteins contribute to the role of human papillomavirus type 16 E7 in the papillomavirus life cycle
Abstract
Human papillomaviruses (HPVs), most commonly the HPV16 genotype, are the principle etiological determinant for cervical cancer, a common cancer worldwide resulting in over 200,000 deaths annually. The oncogenic properties of HPVs are attributable in part to the virally encoded protein E7, best known for its ability to bind to and induce the degradation of the retinoblastoma tumor suppressor, pRb, and related "pocket proteins" p107 and p130. Previously, we defined a role for E7 in the productive stage of the HPV16 life cycle, which takes place in stratified squamous epithelia. HPV perturbs the normal processes of cell growth and differentiation of stratified squamous epithelia. HPVs reprogram cells to support continued DNA synthesis and inhibit their differentiation in the suprabasal compartment of the epithelia, where cells normally have withdrawn from the cell cycle and initiated a well-defined pattern of terminal differentiation. These virus-induced perturbations, which contribute to the production of progeny HPVs, are dependent on E7. In this study, we define the mechanism of action by which E7 contributes to the productive stage of the HPV16 life cycle. We found that the ability of HPV16 to reprogram suprabasal cells to support DNA synthesis correlates with E7's ability to bind pocket proteins but not its ability to induce their degradation. In contrast, the ability of HPV16 to perturb differentiation correlated with both E7's binding to and degradation of pocket proteins. These data indicate that different hallmarks of the productive stage of the HPV16 life cycle rely upon different sets of requirements for E7.
Figures
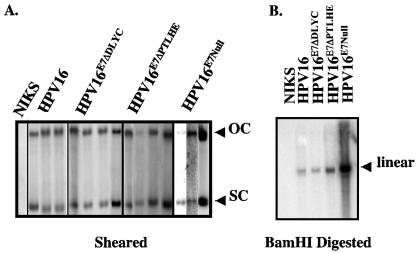
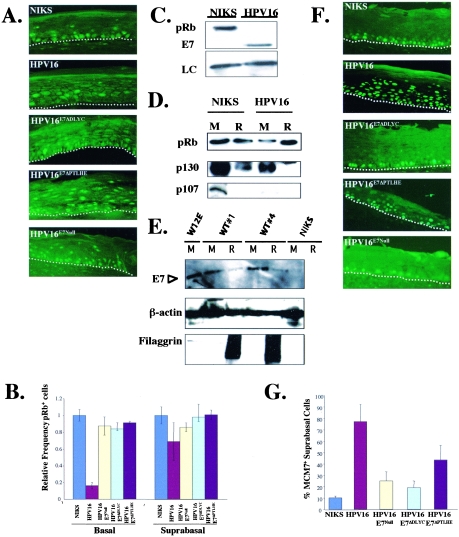
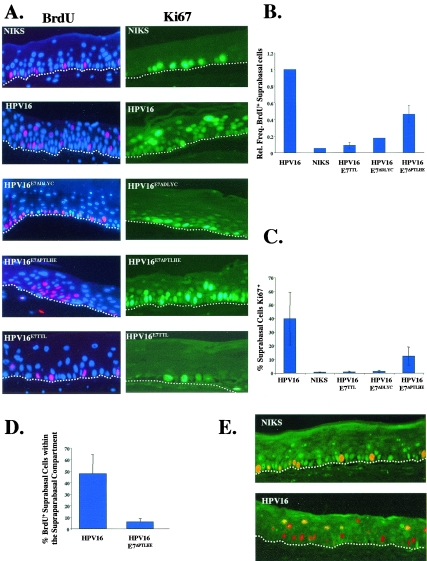
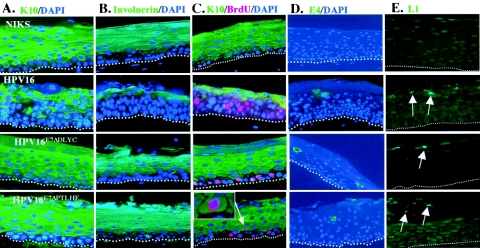
Similar articles
-
Quantitative role of the human papillomavirus type 16 E5 gene during the productive stage of the viral life cycle.J Virol. 2003 Mar;77(5):2832-42. doi: 10.1128/jvi.77.5.2832-2842.2003. J Virol. 2003. PMID: 12584306 Free PMC article.
-
The human papillomavirus type 16 E7 oncogene is required for the productive stage of the viral life cycle.J Virol. 2000 Jul;74(14):6622-31. doi: 10.1128/jvi.74.14.6622-6631.2000. J Virol. 2000. PMID: 10864676 Free PMC article.
-
The E7 proteins of low- and high-risk human papillomaviruses share the ability to target the pRB family member p130 for degradation.Proc Natl Acad Sci U S A. 2006 Jan 10;103(2):437-42. doi: 10.1073/pnas.0510012103. Epub 2005 Dec 28. Proc Natl Acad Sci U S A. 2006. PMID: 16381817 Free PMC article.
-
Cellular and molecular alterations in human epithelial cells transformed by recombinant human papillomavirus DNA.Crit Rev Oncog. 1993;4(4):337-60. Crit Rev Oncog. 1993. PMID: 8394744 Review.
-
Cellular targets of the oncoproteins encoded by the cancer associated human papillomaviruses.Princess Takamatsu Symp. 1991;22:239-48. Princess Takamatsu Symp. 1991. PMID: 1668886 Review.
Cited by
-
The papillomavirus E7 proteins.Virology. 2013 Oct;445(1-2):138-68. doi: 10.1016/j.virol.2013.04.013. Epub 2013 May 31. Virology. 2013. PMID: 23731972 Free PMC article. Review.
-
Cellular transformation by human papillomaviruses: lessons learned by comparing high- and low-risk viruses.Virology. 2012 Mar 15;424(2):77-98. doi: 10.1016/j.virol.2011.12.018. Epub 2012 Jan 27. Virology. 2012. PMID: 22284986 Free PMC article. Review.
-
Metabolic Control by DNA Tumor Virus-Encoded Proteins.Pathogens. 2021 May 6;10(5):560. doi: 10.3390/pathogens10050560. Pathogens. 2021. PMID: 34066504 Free PMC article. Review.
-
Recent advances in the search for antiviral agents against human papillomaviruses.Antivir Ther. 2007;12(4):431-51. Antivir Ther. 2007. PMID: 17668552 Free PMC article. Review.
-
Association between promoter methylation and gene expression of CGB3 and NOP56 in HPV-infected cervical cancer cells.Biomed Rep. 2022 Jan;16(1):1. doi: 10.3892/br.2021.1484. Epub 2021 Nov 3. Biomed Rep. 2022. PMID: 34820124 Free PMC article.
References
-
- Balsitis, S. J., J. Sage, S. Duensing, K. Munger, T. Jacks, and P. F. Lambert. 2003. Recapitulation of the effects of the human papillomavirus type 16 E7 oncogene on mouse epithelium by somatic Rb deletion and detection of pRb-independent effects of E7 in vivo. Mol. Cell. Biol. 23:9094-9103. - PMC - PubMed
-
- Brown, D. C., and K. C. Gatter. 2002. Ki67 protein: the immaculate deception? Histopathology 40:2-11. - PubMed
-
- Cam, H., and B. D. Dynlacht. 2003. Emerging roles for E2F: beyond the G1/S transition and DNA replication. Cancer Cell 3:311-316. - PubMed
-
- Chen, P. L., D. J. Riley, Y. Chen, and W. H. Lee. 1996. Retinoblastoma protein positively regulates terminal adipocyte differentiation through direct interaction with C/EBPs. Genes Dev. 10:2794-2804. - PubMed
-
- Chien, W. M., J. N. Parker, D. C. Schmidt-Grimminger, T. R. Broker, and L. T. Chow. 2000. Casein kinase II phosphorylation of the human papillomavirus-18 E7 protein is critical for promoting S-phase entry. Cell Growth Differ. 11:425-435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
