

Characterization of adeno-associated virus genomes isolated from human tissues
- PMID: 16282479
- PMCID: PMC1287572
- DOI: 10.1128/JVI.79.23.14793-14803.2005
Characterization of adeno-associated virus genomes isolated from human tissues
Abstract
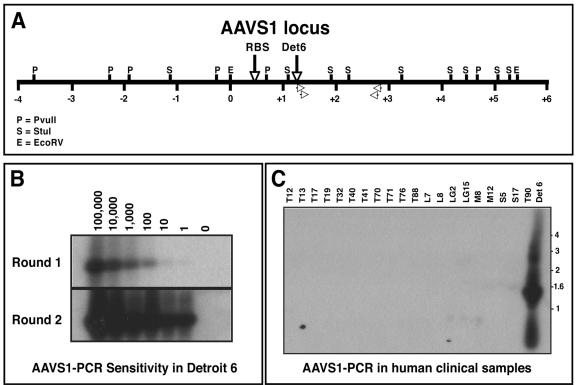
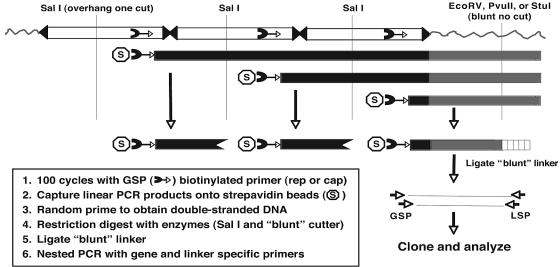
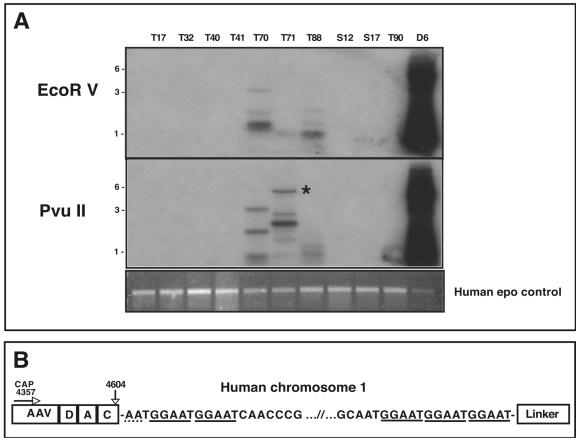
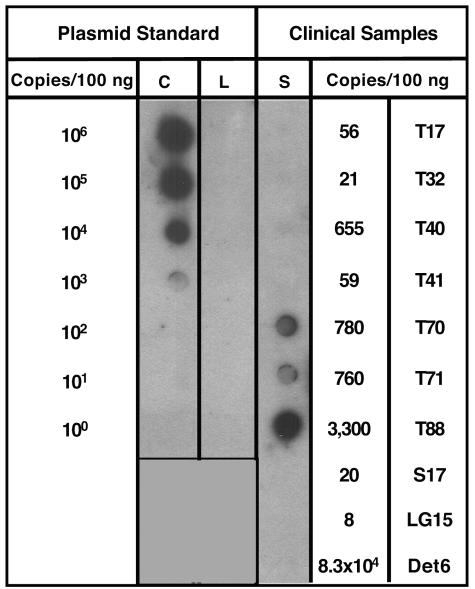
Infection with wild-type adeno-associated virus (AAV) is common in humans, but very little is known about the in vivo biology of AAV. On a molecular level, it has been shown in cultured cells that AAV integrates in a site-specific manner on human chromosome 19, but this has never been demonstrated directly in infected human tissues. To that end, we tested 175 tissue samples for the presence of AAV DNA, and when present, examined the specific form of the viral DNA. AAV was detected in 7 of 101 tonsil-adenoid samples and in 2 of 74 other tissue samples (spleen and lung). In these nine samples, we were unable to detect AAV integration in the AAVS1 locus using a sensitive PCR assay designed to amplify specific viral-cellular DNA junctions. Additionally, we used a second complementary assay, linear amplification-mediated-PCR (LAM-PCR) to widen our search for integration events. Analysis of individual LAM-PCR products revealed that the AAV genomes were arranged predominantly in a head-to-tail array, with deletions and extensive rearrangements in the inverted terminal repeat sequences. A single AAV-cellular junction was identified from a tonsil sample and it mapped to a highly repetitive satellite DNA element on chromosome 1. Given these data, we entertained the possibility that instead of integrated forms, AAV genomes were present as extrachromosomal forms. We used a novel amplification assay (linear rolling-circle amplification) to show that the majority of wild-type AAV DNA existed as circular double-stranded episomes in our tissues. Thus, following naturally acquired infection, AAV DNA can persist mainly as circular episomes in human tissues. These findings are consistent with the circular episomal forms of recombinant AAV vectors that have been isolated and characterized from in vivo transduced tissues.
Figures



Similar articles
-
Cellular recombination pathways and viral terminal repeat hairpin structures are sufficient for adeno-associated virus integration in vivo and in vitro.J Virol. 1997 Dec;71(12):9231-47. doi: 10.1128/JVI.71.12.9231-9247.1997. J Virol. 1997. PMID: 9371582 Free PMC article.
-
Adeno-associated virus (AAV) site-specific recombination does not require a Rep-dependent origin of replication within the AAV terminal repeat.Proc Natl Acad Sci U S A. 2001 Nov 20;98(24):13525-30. doi: 10.1073/pnas.241508998. Epub 2001 Nov 13. Proc Natl Acad Sci U S A. 2001. PMID: 11707592 Free PMC article.
-
Molecular characterization of adeno-associated viruses infecting children.J Virol. 2005 Dec;79(23):14781-92. doi: 10.1128/JVI.79.23.14781-14792.2005. J Virol. 2005. PMID: 16282478 Free PMC article.
-
Integration of adeno-associated virus (AAV) and recombinant AAV vectors.Annu Rev Genet. 2004;38:819-45. doi: 10.1146/annurev.genet.37.110801.143717. Annu Rev Genet. 2004. PMID: 15568995 Review.
-
Site-specific integration by adeno-associated virus.Proc Natl Acad Sci U S A. 1996 Oct 15;93(21):11288-94. doi: 10.1073/pnas.93.21.11288. Proc Natl Acad Sci U S A. 1996. PMID: 8876128 Free PMC article. Review.
Cited by
-
Intra-tracheal delivery of AAV6 vectors results in sustained transduction in murine lungs without genomic integration.Gene X. 2020 Jul 31;5:100037. doi: 10.1016/j.gene.2020.100037. eCollection 2020 Dec. Gene X. 2020. PMID: 32904225 Free PMC article.
-
Adeno-associated virus vector as a platform for gene therapy delivery.Nat Rev Drug Discov. 2019 May;18(5):358-378. doi: 10.1038/s41573-019-0012-9. Nat Rev Drug Discov. 2019. PMID: 30710128 Free PMC article. Review.
-
Identification of Extrachromosomal Linear microDNAs Interacted with microRNAs in the Cell Nuclei.Cells. 2019 Feb 1;8(2):111. doi: 10.3390/cells8020111. Cells. 2019. PMID: 30717295 Free PMC article.
-
Life-Long AAV-Mediated CRISPR Genome Editing in Dystrophic Heart Improves Cardiomyopathy without Causing Serious Lesions in mdx Mice.Mol Ther. 2019 Aug 7;27(8):1407-1414. doi: 10.1016/j.ymthe.2019.05.001. Epub 2019 May 15. Mol Ther. 2019. PMID: 31129119 Free PMC article.
-
Tetramer-Based Enrichment of Preexisting Anti-AAV8 CD8+ T Cells in Human Donors Allows the Detection of a TEMRA Subpopulation.Front Immunol. 2020 Jan 21;10:3110. doi: 10.3389/fimmu.2019.03110. eCollection 2019. Front Immunol. 2020. PMID: 32038634 Free PMC article.
References
-
- Atchison, R. W., B. C. Casto, and W. M. Hammon. 1965. Adenovirus-associated defective virus particles. Science 149:754-756. - PubMed
-
- Atchison, R. W., B. C. Casto, and W. M. Hammon. 1966. Electron microscopy of adenovirus-associated virus (AAV) in cell cultures. Virology 29:353-357. - PubMed
-
- Berns, K. I., T. C. Pinkerton, G. F. Thomas, and M. D. Hoggan. 1975. Detection of adeno-associated virus (AAV)-specific nucleotide sequences in DNA isolated from latently infected Detroit 6 cells. Virology 68:556-560. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
