

Avidity for antigen shapes clonal dominance in CD8+ T cell populations specific for persistent DNA viruses
- PMID: 16287711
- PMCID: PMC2212993
- DOI: 10.1084/jem.20051357
Avidity for antigen shapes clonal dominance in CD8+ T cell populations specific for persistent DNA viruses
Abstract
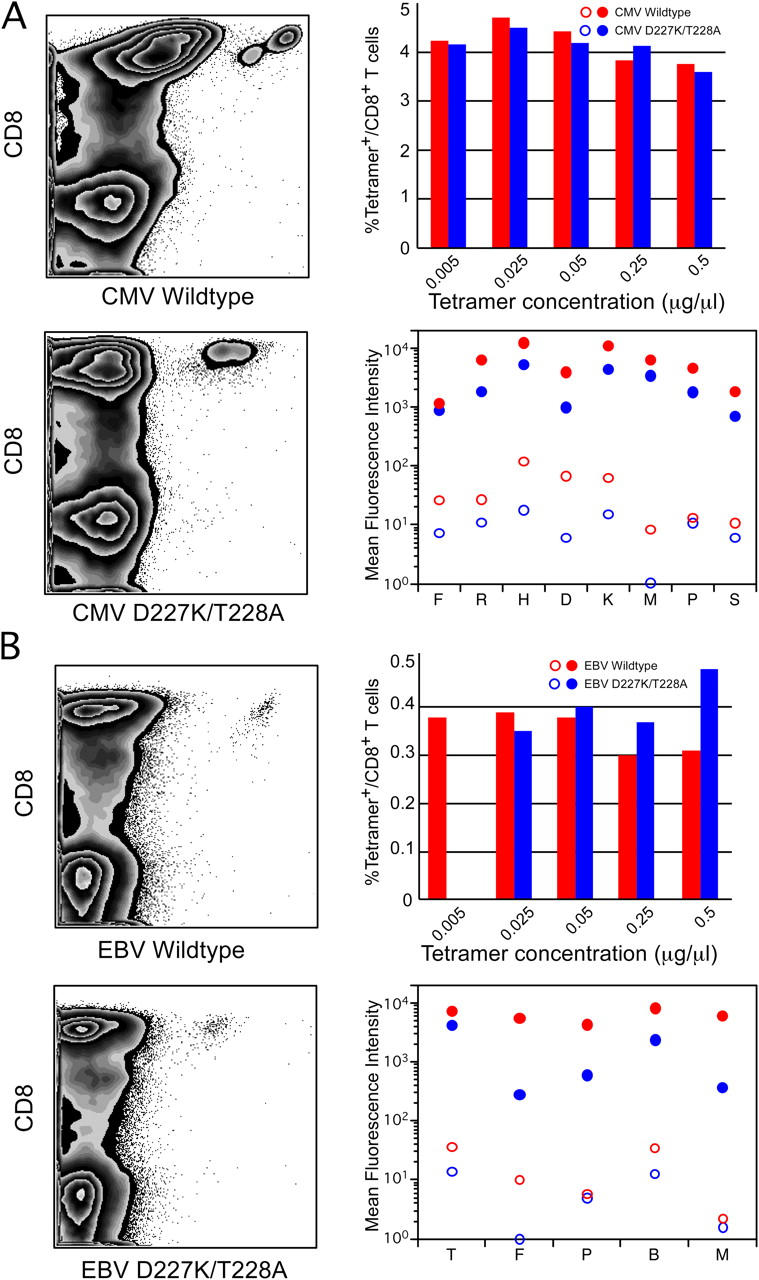
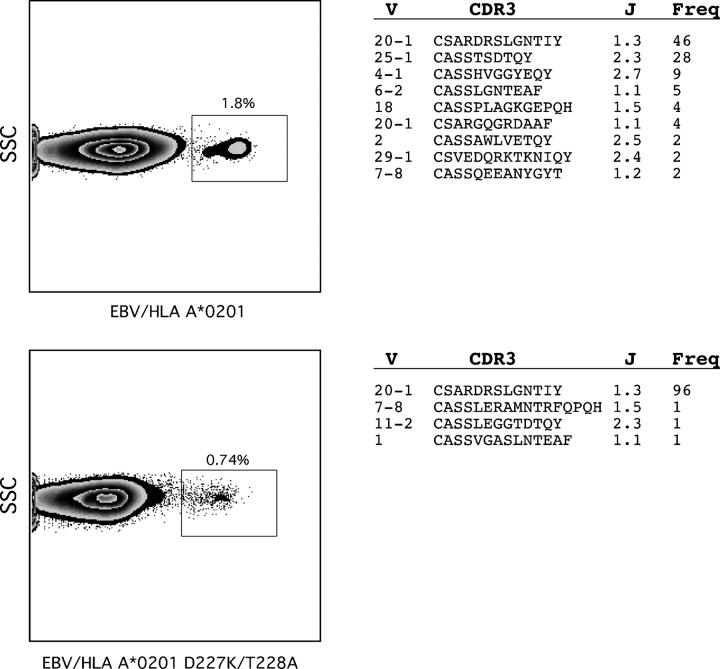
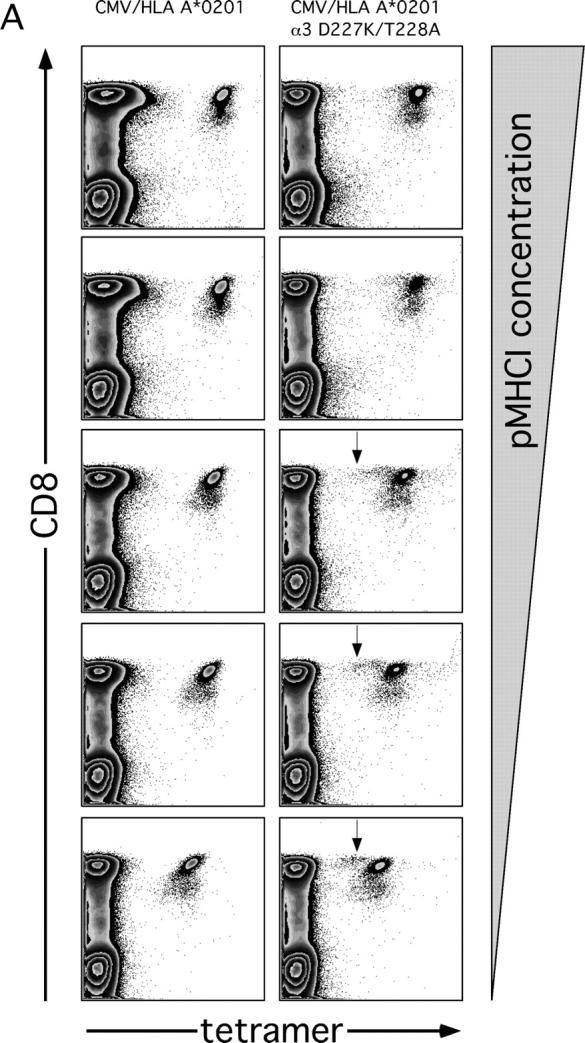
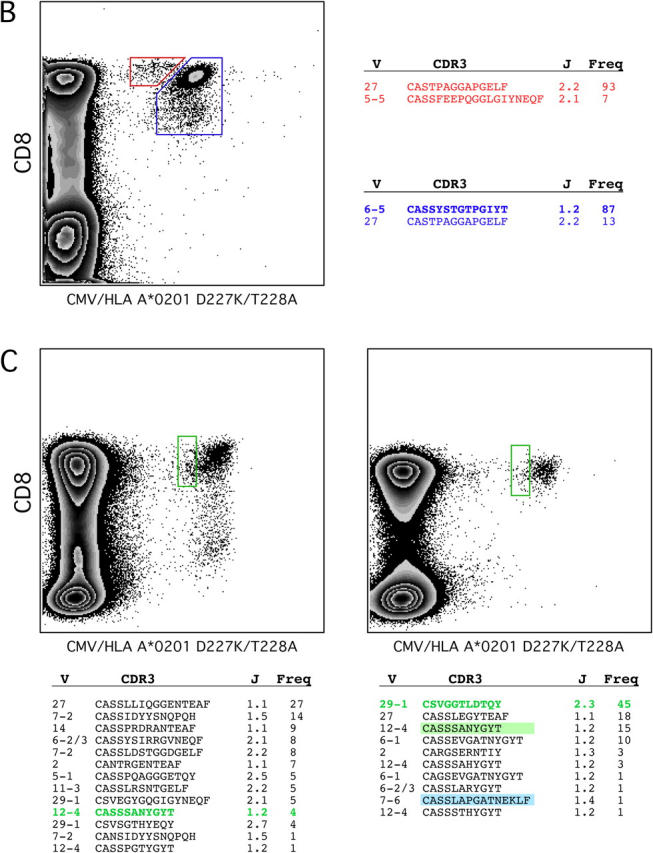
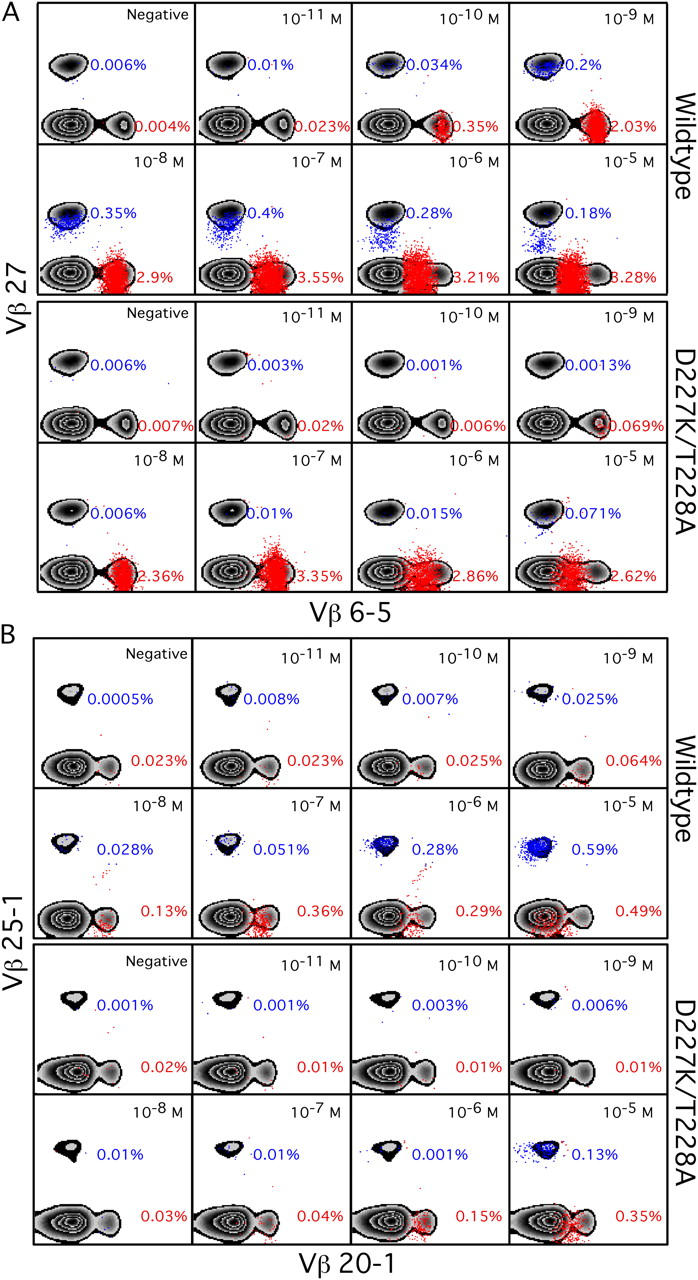
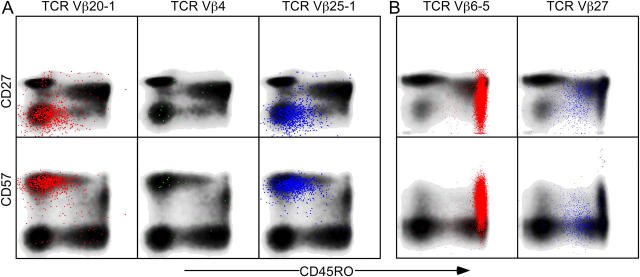
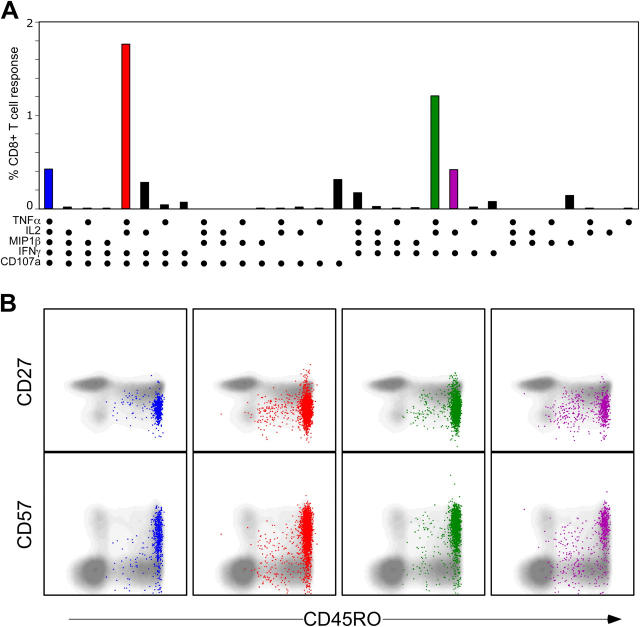
The forces that govern clonal selection during the genesis and maintenance of specific T cell responses are complex, but amenable to decryption by interrogation of constituent clonotypes within the antigen-experienced T cell pools. Here, we used point-mutated peptide-major histocompatibility complex class I (pMHCI) antigens, unbiased TCRB gene usage analysis, and polychromatic flow cytometry to probe directly ex vivo the clonal architecture of antigen-specific CD8(+) T cell populations under conditions of persistent exposure to structurally stable virus-derived epitopes. During chronic infection with cytomegalovirus and Epstein-Barr virus, CD8(+) T cell responses to immunodominant viral antigens were oligoclonal, highly skewed, and exhibited diverse clonotypic configurations; TCRB CDR3 sequence analysis indicated positive selection at the protein level. Dominant clonotypes demonstrated high intrinsic antigen avidity, defined strictly as a physical parameter, and were preferentially driven toward terminal differentiation in phenotypically heterogeneous populations. In contrast, subdominant clonotypes were characterized by lower intrinsic avidities and proportionately greater dependency on the pMHCI-CD8 interaction for antigen uptake and functional sensitivity. These findings provide evidence that interclonal competition for antigen operates in human T cell populations, while preferential CD8 coreceptor compensation mitigates this process to maintain clonotypic diversity. Vaccine strategies that reconstruct these biological processes could generate T cell populations that mediate optimal delivery of antiviral effector function.
Figures

Similar articles
-
Clonotype selection and composition of human CD8 T cells specific for persistent herpes viruses varies with differentiation but is stable over time.J Immunol. 2009 Jul 1;183(1):319-31. doi: 10.4049/jimmunol.0803647. J Immunol. 2009. PMID: 19542443
-
The TCR repertoire of an immunodominant CD8+ T lymphocyte population.J Immunol. 2001 Apr 1;166(7):4525-33. doi: 10.4049/jimmunol.166.7.4525. J Immunol. 2001. PMID: 11254709
-
Mutations in a dominant Nef epitope of simian immunodeficiency virus diminish TCR:epitope peptide affinity but not epitope peptide:MHC class I binding.J Immunol. 2011 Sep 15;187(6):3300-13. doi: 10.4049/jimmunol.1101080. Epub 2011 Aug 12. J Immunol. 2011. PMID: 21841125 Free PMC article.
-
Tracking phenotypically and functionally distinct T cell subsets via T cell repertoire diversity.Mol Immunol. 2008 Feb;45(3):607-18. doi: 10.1016/j.molimm.2006.05.017. Epub 2007 Aug 24. Mol Immunol. 2008. PMID: 17719639 Free PMC article. Review.
-
Techniques to improve the direct ex vivo detection of low frequency antigen-specific CD8+ T cells with peptide-major histocompatibility complex class I tetramers.Cytometry A. 2008 Nov;73(11):1001-9. doi: 10.1002/cyto.a.20642. Cytometry A. 2008. PMID: 18836993 Free PMC article. Review.
Cited by
-
Genetic and structural basis for selection of a ubiquitous T cell receptor deployed in Epstein-Barr virus infection.PLoS Pathog. 2010 Nov 18;6(11):e1001198. doi: 10.1371/journal.ppat.1001198. PLoS Pathog. 2010. PMID: 21124993 Free PMC article.
-
The Memory-CD8+-T-Cell Response to Conserved Influenza Virus Epitopes in Mice Is Not Influenced by Time Since Previous Infection.Vaccines (Basel). 2024 Apr 15;12(4):419. doi: 10.3390/vaccines12040419. Vaccines (Basel). 2024. PMID: 38675801 Free PMC article.
-
A T Cell Receptor Locus Harbors a Malaria-Specific Immune Response Gene.Immunity. 2017 Nov 21;47(5):835-847.e4. doi: 10.1016/j.immuni.2017.10.013. Epub 2017 Nov 14. Immunity. 2017. PMID: 29150238 Free PMC article.
-
Combined immunodeficiency and Epstein-Barr virus-induced B cell malignancy in humans with inherited CD70 deficiency.J Exp Med. 2017 Jan;214(1):91-106. doi: 10.1084/jem.20160849. Epub 2016 Dec 23. J Exp Med. 2017. PMID: 28011864 Free PMC article.
-
A Unique T-Cell Receptor Amino Acid Sequence Selected by Human T-Cell Lymphotropic Virus Type 1 Tax301-309-Specific Cytotoxic T Cells in HLA-A24:02-Positive Asymptomatic Carriers and Adult T-Cell Leukemia/Lymphoma Patients.J Virol. 2017 Sep 12;91(19):e00974-17. doi: 10.1128/JVI.00974-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28724766 Free PMC article.
References
-
- van der Merwe, P.A., and S.J. Davis. 2003. Molecular interactions mediating T cell antigen recognition. Annu. Rev. Immunol. 21:659–684. - PubMed
-
- Kedl, R.M., J.W. Kappler, and P. Marrack. 2003. Epitope dominance, competition and T cell affinity maturation. Curr. Opin. Immunol. 15:120–127. - PubMed
-
- Smith, A.L., M.E. Wikstrom, and B. Fazekas de St Groth. 2000. Visualizing T cell competition for peptide/MHC complexes: a specific mechanism to minimize the effect of precursor frequency. Immunity. 13:783–794. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
