

The role of cuticular pheromones in courtship conditioning of Drosophila males
- PMID: 16287720
- PMCID: PMC1356183
- DOI: 10.1101/lm.85605
The role of cuticular pheromones in courtship conditioning of Drosophila males
Abstract
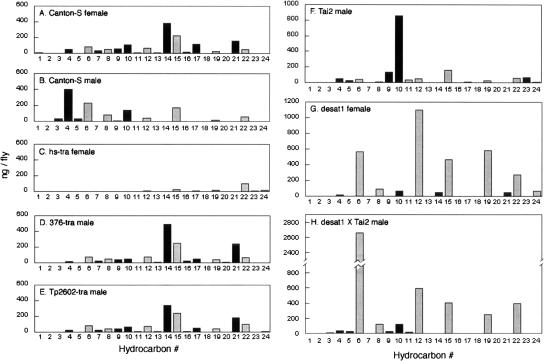
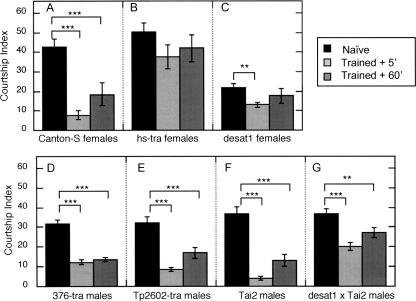
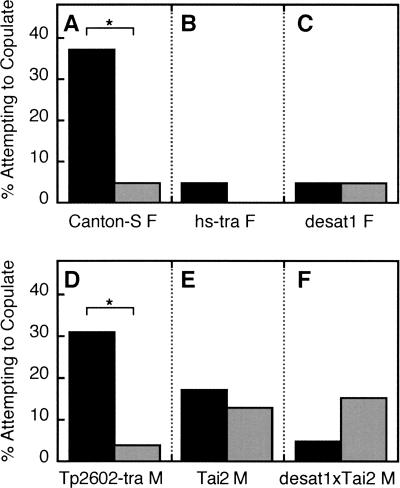
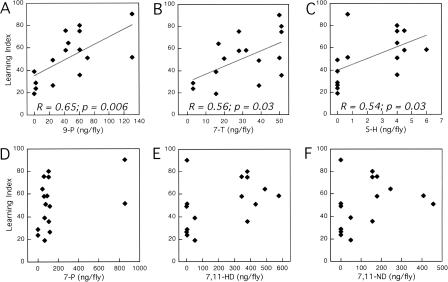
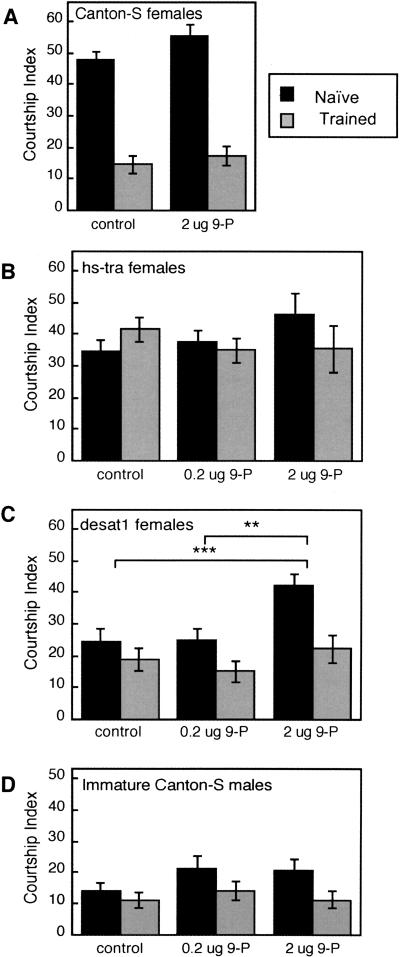
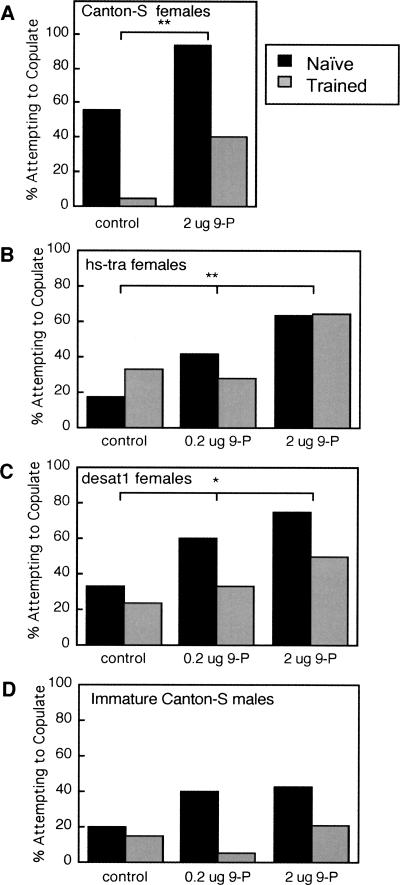
Courtship conditioning is an associative learning paradigm in Drosophila melanogaster, wherein male courtship behavior is modified by experience with unreceptive, previously mated females. While the training experience with mated females involves multiple sensory and behavioral interactions, we hypothesized that female cuticular hydrocarbons function as a specific chemosensory conditioned stimulus in this learning paradigm. The effects of training with mated females were determined in courtship tests with either wild-type virgin females as courtship targets, or with target flies of different genotypes that express distinct cuticular hydrocarbon (CH) profiles. Results of tests with female targets that lacked the normal CH profile, and with male targets that expressed typically female CH profiles, indicated that components of this CH profile are both necessary and sufficient cues to elicit the effects of conditioning. Results with additional targets indicated that the female-specific 7,11-dienes, which induce naive males to court, are not essential components of the conditioned stimulus. Rather, the learned response was significantly correlated with the levels of 9-pentacosene (9-P), a compound found in both males and females of many Drosophila strains and species. Adding 9-P to target flies showed that it stimulates courting males to attempt to copulate, and confirmed its role as a component of the conditioned stimulus by demonstrating dose-dependent increases in the expression of the learned response. Thus, 9-P can contribute significantly to the conditioned suppression of male courtship toward targets that express this pheromone.
Figures
Similar articles
-
Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate.Curr Biol. 2007 Apr 3;17(7):599-605. doi: 10.1016/j.cub.2007.01.053. Epub 2007 Mar 15. Curr Biol. 2007. PMID: 17363250 Free PMC article.
-
Courtship Conditioning/Suppression Assays in Drosophila.Cold Spring Harb Protoc. 2023 Jul 5;2023(7):.pdb.prot108106. doi: 10.1101/pdb.prot108106. Cold Spring Harb Protoc. 2023. PMID: 36781210
-
Contribution of oenocytes and pheromones to courtship behaviour in Drosophila.BMC Biochem. 2009 Aug 11;10:21. doi: 10.1186/1471-2091-10-21. BMC Biochem. 2009. PMID: 19671131 Free PMC article.
-
Drosophila conditioned courtship: two ways of testing memory.Learn Mem. 1999 Jan-Feb;6(1):1-20. Learn Mem. 1999. PMID: 10355520 Free PMC article. Review.
-
Conditioned courtship suppression in Drosophila melanogaster.J Neurogenet. 2021 Sep;35(3):154-167. doi: 10.1080/01677063.2021.1873323. Epub 2021 Jan 31. J Neurogenet. 2021. PMID: 33522326 Review.
Cited by
-
A pleiotropic chemoreceptor facilitates the production and perception of mating pheromones.iScience. 2022 Dec 29;26(1):105882. doi: 10.1016/j.isci.2022.105882. eCollection 2023 Jan 20. iScience. 2022. PMID: 36691619 Free PMC article.
-
Evolution of Reproductive Behavior.Genetics. 2020 Jan;214(1):49-73. doi: 10.1534/genetics.119.302263. Genetics. 2020. PMID: 31907301 Free PMC article. Review.
-
Characterization of a Drosophila Alzheimer's disease model: pharmacological rescue of cognitive defects.PLoS One. 2011;6(6):e20799. doi: 10.1371/journal.pone.0020799. Epub 2011 Jun 6. PLoS One. 2011. PMID: 21673973 Free PMC article.
-
Drosophila cuticular hydrocarbons revisited: mating status alters cuticular profiles.PLoS One. 2010 Mar 9;5(3):e9607. doi: 10.1371/journal.pone.0009607. PLoS One. 2010. PMID: 20231905 Free PMC article.
-
Natural variation in the strength and direction of male mating preferences for female pheromones in Drosophila melanogaster.PLoS One. 2014 Jan 28;9(1):e87509. doi: 10.1371/journal.pone.0087509. eCollection 2014. PLoS One. 2014. PMID: 24489930 Free PMC article.
References
-
- Ackerman, S.L. and Siegel, R.W. 1986. Chemically reinforced conditioned courtship in Drosophila: Responses of wild-type and the dunce, amnesiac, and don giovanni mutants. J. Neurogenet. 3: 111-123. - PubMed
-
- Amrein, H. 2004. Pheromone perception and behavior in Drosophila. Curr. Opin. Neurobiol. 14: 435-442. - PubMed
-
- Antony, C. and Jallon, J.-M. 1982. The chemical basis for sex recognition in Drosophila melanogaster. J. Insect Physiol. 28: 873-880.
-
- Antony, C., Davis, T.L., Carlson, D.A., Pechine, J.-M., and Jallon, J.-M. 1985. Compared behavioral responses of male Drosophila melanogaster (Canton S) to natural and synthetic aphrodisiacs. J. Chem. Ecol. 11: 1617-1629. - PubMed
-
- Bastock, M. and Manning, A. 1955. The courtship of Drosophila melanogaster. Behaviour 8: 85-111.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases