

Minimal functions and physiological conditions required for growth of salmonella enterica on ethanolamine in the absence of the metabolosome
- PMID: 16291677
- PMCID: PMC1291257
- DOI: 10.1128/JB.187.23.8039-8046.2005
Minimal functions and physiological conditions required for growth of salmonella enterica on ethanolamine in the absence of the metabolosome
Abstract
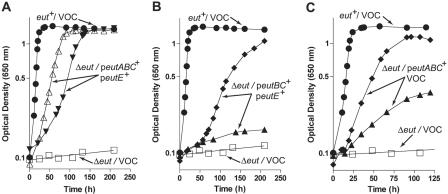
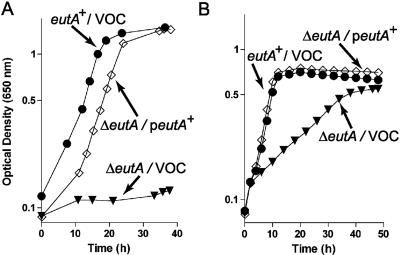
During growth on ethanolamine, Salmonella enterica synthesizes a multimolecular structure that mimics the carboxysome used by some photosynthetic bacteria to fix CO(2). In S. enterica, this carboxysome-like structure (hereafter referred to as the ethanolamine metabolosome) is thought to contain the enzymatic machinery needed to metabolize ethanolamine into acetyl coenzyme A (acetyl-CoA). Analysis of the growth behavior of mutant strains of S. enterica lacking specific functions encoded by the 17-gene ethanolamine utilization (eut) operon established the minimal biochemical functions needed by this bacterium to use ethanolamine as a source of carbon and energy. The data obtained support the conclusion that the ethanolamine ammnonia-lyase (EAL) enzyme (encoded by the eutBC genes) and coenzyme B(12) are necessary and sufficient to grow on ethanolamine. We propose that the EutD phosphotransacetylase and EutG alcohol dehydrogenase are important to maintain metabolic balance. Glutathione (GSH) had a strong positive effect that compensated for the lack of the EAL reactivase EutA protein under aerobic growth on ethanolamine. Neither GSH nor EutA was needed during growth on ethanolamine under reduced-oxygen conditions. GSH also stimulated growth of a strain lacking the acetaldehyde dehydrogenase (EutE) enzyme. The role of GSH in ethanolamine catabolism is complex and requires further investigation. Our data show that the ethanolamine metabolosome is not involved in the biochemistry of ethanolamine catabolism. We propose the metabolosome is needed to concentrate low levels of ethanolamine catabolic enzymes, to keep the level of toxic acetaldehyde low, to generate enough acetyl-CoA to support cell growth, and to maintain a pool of free CoA.
Figures




References
-
- Abend, A., V. Bandarian, R. Nitsche, E. Stupperich, J. Retey, and G. H. Reed. 1999. Ethanolamine ammonia-lyase has a “base-on” binding mode for coenzyme B12. Arch. Biochem. Biophys. 370:138-141. - PubMed
-
- Badger, M. R., and G. D. Price. 2003. CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J. Exp. Bot. 54:609-622. - PubMed
-
- Bobik, T. A., G. D. Havemann, R. J. Busch, D. S. Williams, and H. C. Aldrich. 1999. The propanediol utilization (pdu) operon of Salmonella enterica serovar Typhimurium LT2 includes genes necessary for formation of polyhedral organelles involved in coenzyme B12-dependent 1,2-propanediol degradation. J. Bacteriol. 181:5967-5975. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
