

Age-dependent neurofibrillary tangle formation, neuron loss, and memory impairment in a mouse model of human tauopathy (P301L)
- PMID: 16291936
- PMCID: PMC6725849
- DOI: 10.1523/JNEUROSCI.3279-05.2005
Age-dependent neurofibrillary tangle formation, neuron loss, and memory impairment in a mouse model of human tauopathy (P301L)
Abstract
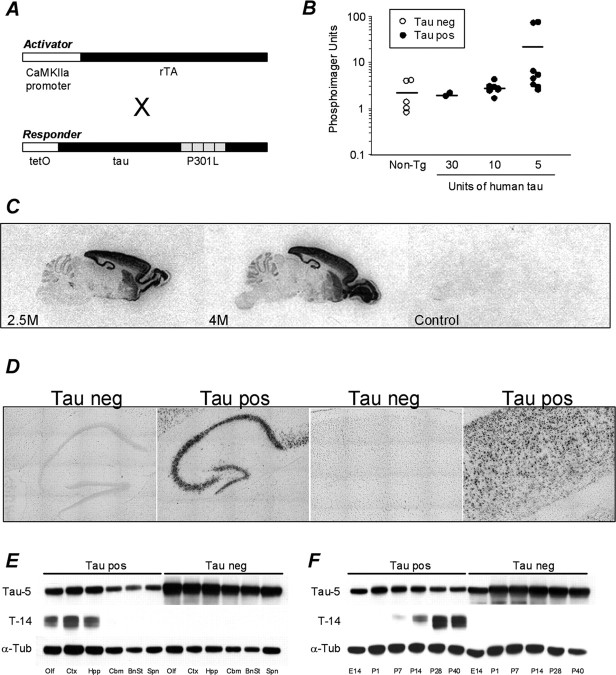
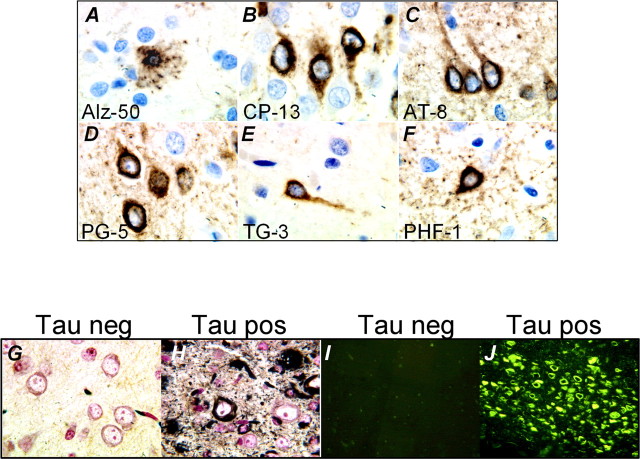
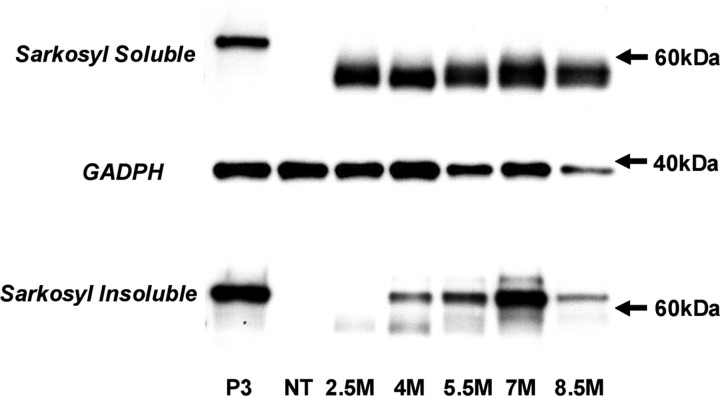
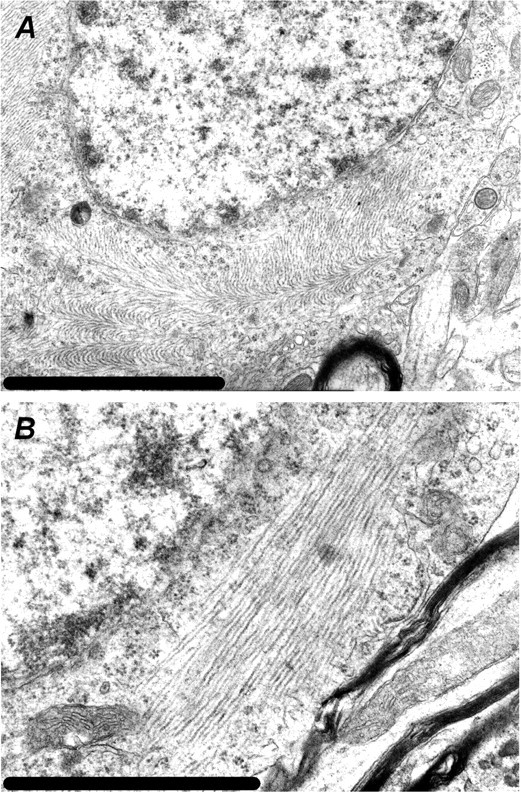
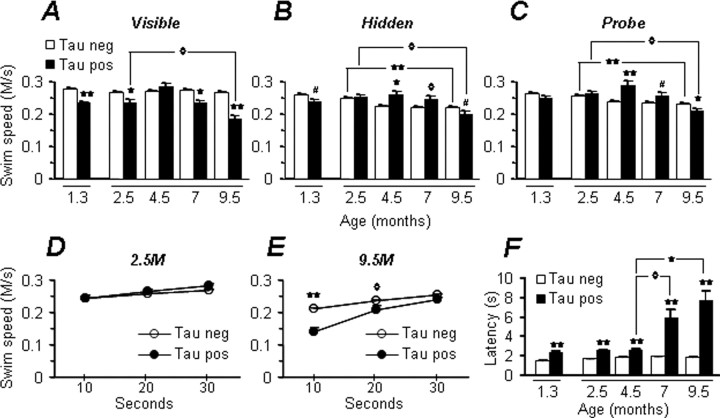
Here, we describe the generation of a novel transgenic mouse model of human tauopathy. The rTg(tau(P301L))4510 mouse expresses the P301L mutation in tau (4R0N) associated with frontotemporal dementia and parkinsonism linked to chromosome 17. Transgene expression was driven by a forebrain-specific Ca(2+) calmodulin kinase II promoter system resulting in high levels of expression in the hippocampus and neocortex. Importantly, transgene expression in this model is induced via the tetracycline-operon responsive element and is suppressed after treatment with doxycycline. Continued transgene expression in rTg(tau(P301L))4510 mice results in age-dependent development of many salient characteristics of hereditary human dementia. From an early age, immunohistochemical studies demonstrated abnormal biochemical processing of tau and the presence of pathological conformation- and phosphorylation-dependent epitopes. Neurofibrillary tangle (NFT) pathology was first observed in the neocortex and progressed into the hippocampus and limbic structures with increasing age. Consistent with the formation of NFTs, immunoblots indicated an age-dependent transition of accumulating tau species from Sarkosyl soluble 55 kDa to insoluble hyperphosphorylated 64 kDa. Ultrastructural analysis revealed the presence of straight tau filaments. Furthermore, the effects of tau(P301L) expression on spatial reference memory were longitudinally tested using the Morris water maze. Compared with nontransgenic age-matched control littermates, rTg(tau(P301L))4510 mice developed significant cognitive impairments from 4 months of age. Memory deficits were accompanied by gross forebrain atrophy and a prominent loss of neurons, most strikingly in hippocampal subdivision CA1. Collectively, these data describe a novel transgenic mouse that closely mimics human tauopathy and may represent an important model for the future study of tau-related neurodegenerative disease.
Figures





References
-
- Allen B, Ingram E, Takao M, Smith MJ, Jakes R, Virdee K, Yoshida H, Holzer M, Craxton M, Emson PC, Atzori C, Migheli A, Crowther RA, Ghetti B, Spillantini MG, Goedert M (2002) Abundant tau filaments and non-apoptotic neurodegeneration in transgenic mice expressing human P301S tau protein. J Neurosci 22: 9340-9351. - PMC - PubMed
-
- Andorfer C, Kress Y, Espinoza M, de Silva R, Tucker KL, Barde YA, Duff K, Davies P (2003) Hyperphosphorylation and aggregation of tau in mice expressing normal human tau isoforms. J Neurochem 86: 582-590. - PubMed
-
- Borchelt DR, Davis J, Fischer M, Lee MK, Slunt HH, Ratovitsky T, Regard J, Copeland NG, Jenkins NA, Sisodia SS, Price DL (1996) A vector for expressing foreign genes in the brains and hearts of transgenic mice. Genet Anal 13: 159-163. - PubMed
-
- Buee L, Bussiere T, Buee-Scherrer V, Delacourte A, Hof PR (2000) Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev 33: 95-130. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous