

The regulation of mDia1 by autoinhibition and its release by Rho*GTP
- PMID: 16292343
- PMCID: PMC1356318
- DOI: 10.1038/sj.emboj.7600879
The regulation of mDia1 by autoinhibition and its release by Rho*GTP
Abstract
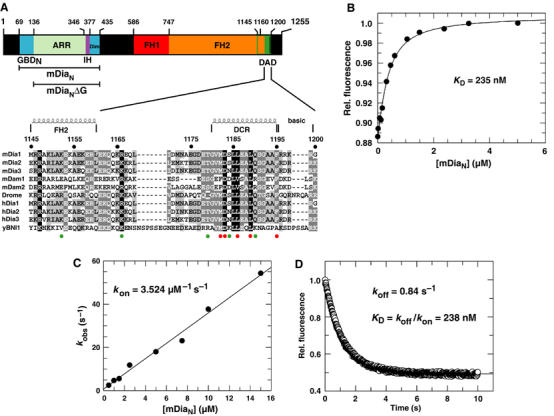
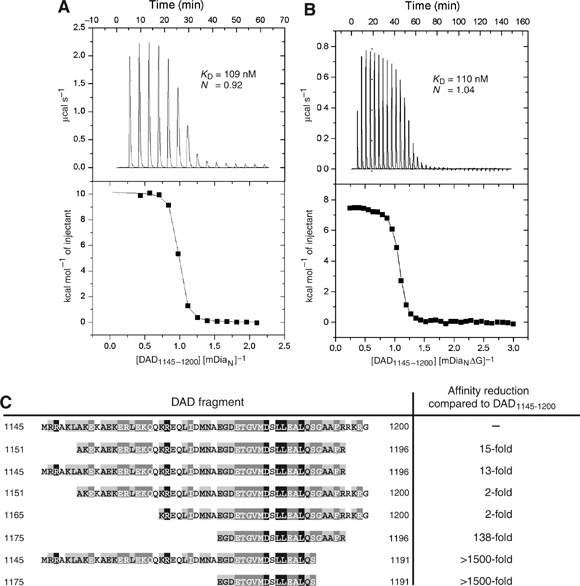
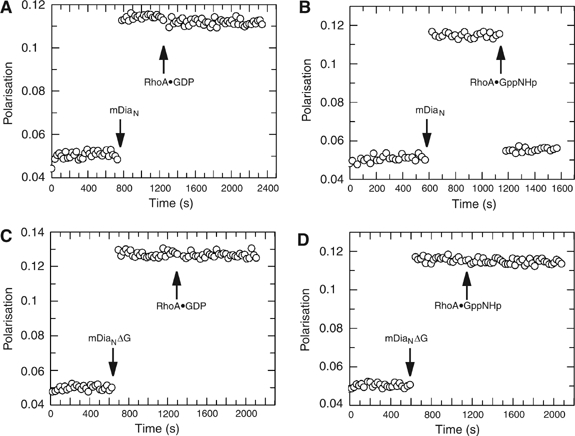
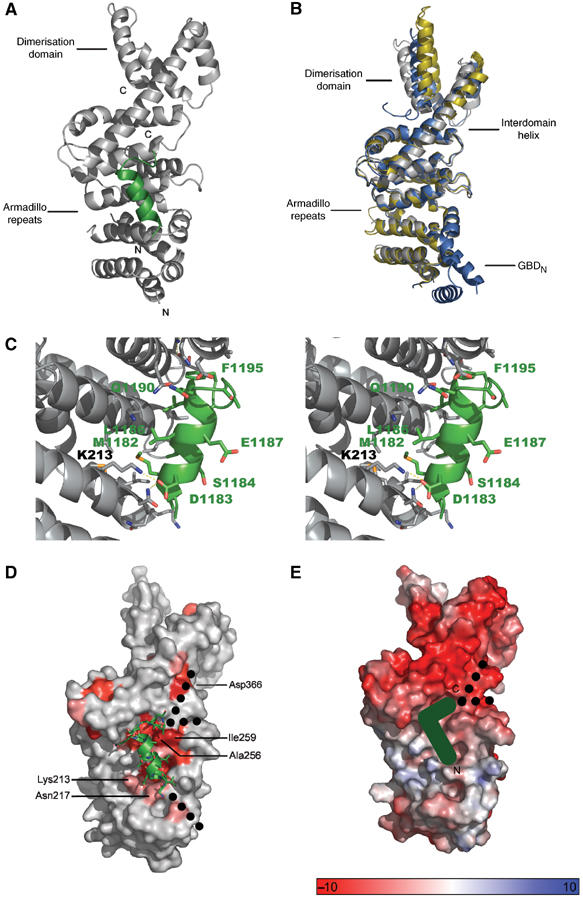
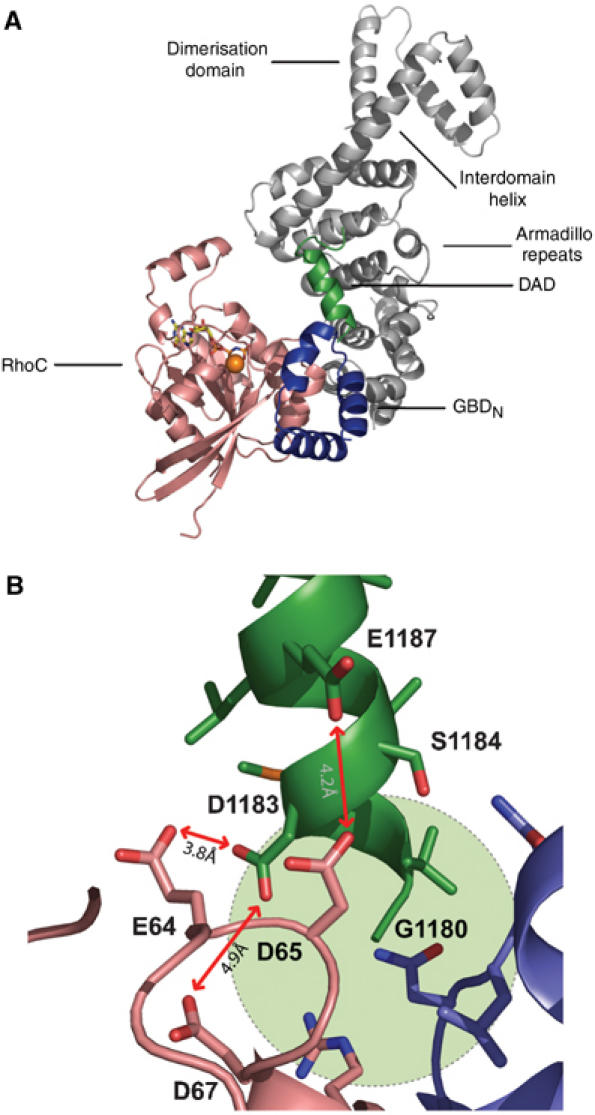
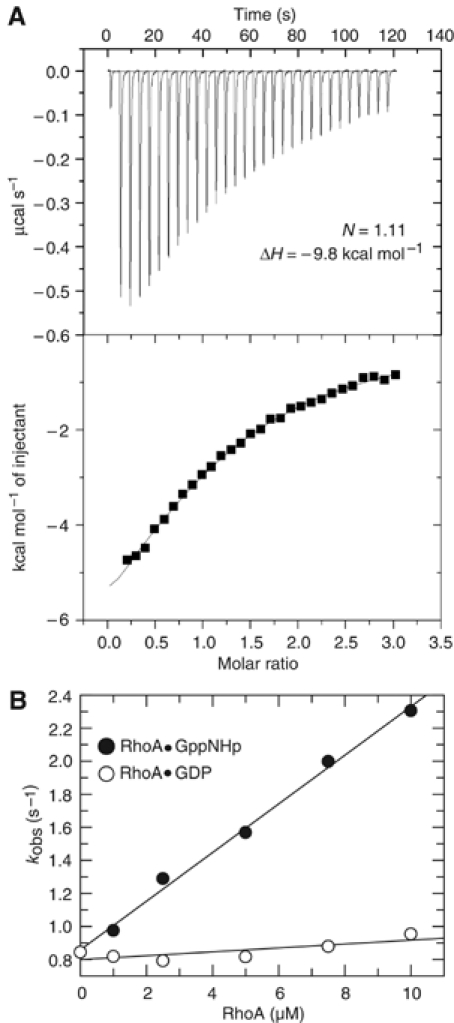
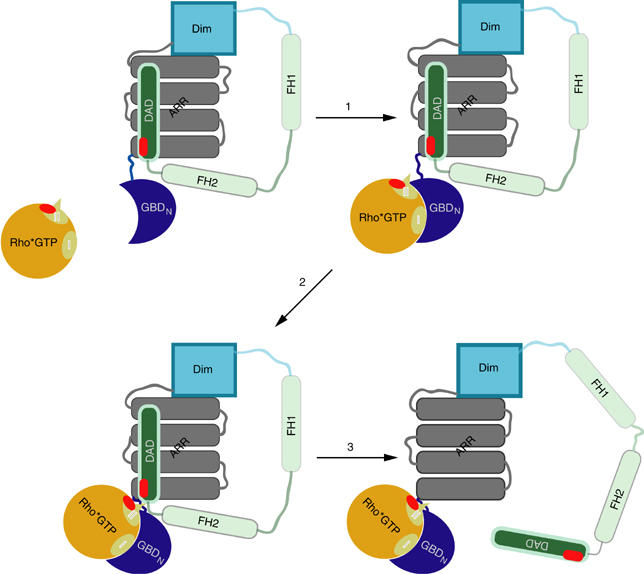
Formins induce the nucleation and polymerisation of unbranched actin filaments via the formin-homology domains 1 and 2. Diaphanous-related formins (Drfs) are regulated by a RhoGTPase-binding domain situated in the amino-terminal (N-terminal) region and a carboxy-terminal Diaphanous-autoregulatory domain (DAD), whose interaction stabilises an autoinhibited inactive conformation. Binding of active Rho releases DAD and activates the catalytic activity of mDia. Here, we report on the interaction of DAD with the regulatory N-terminus of mDia1 (mDia(N)) and its release by Rho*GTP. We have defined the elements required for tight binding and solved the three-dimensional structure of a complex between an mDia(N) construct and DAD by X-ray crystallography. The core DAD region is an alpha-helical peptide, which binds in the most highly conserved region of mDia(N) using mainly hydrophobic interactions. The structure suggests a two-step mechanism for release of autoinhibition whereby Rho*GTP, although having a partially nonoverlapping binding site, displaces DAD by ionic repulsion and steric clashes. We show that Rho*GTP accelerates the dissociation of DAD from the mDia(N)*DAD complex.
Figures
References
-
- Alberts AS (2001) Identification of a carboxyl-terminal Diaphanous-related formin homology protein autoregulatory domain. J Biol Chem 276: 2824–2830 - PubMed
-
- Buck M, Xu W, Rosen MK (2004) A two-state allosteric model for autoinhibition rationalizes WASP signal integration and targeting. J Mol Biol 338: 271–285 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
