

Effects of brefeldin A on pollen germination and tube growth. Antagonistic effects on endocytosis and secretion
- PMID: 16299176
- PMCID: PMC1310552
- DOI: 10.1104/pp.105.069765
Effects of brefeldin A on pollen germination and tube growth. Antagonistic effects on endocytosis and secretion
Abstract
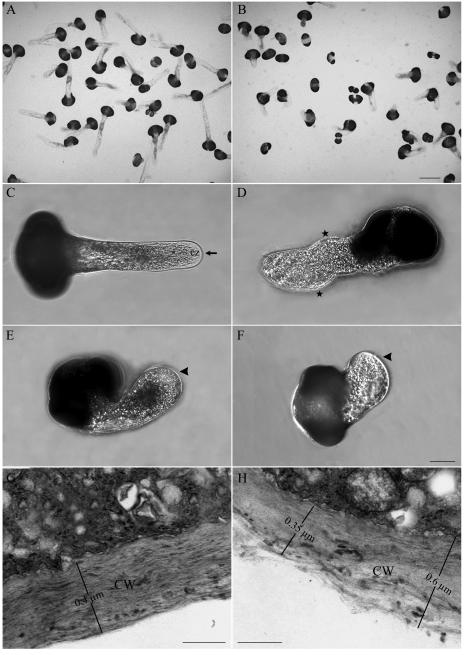
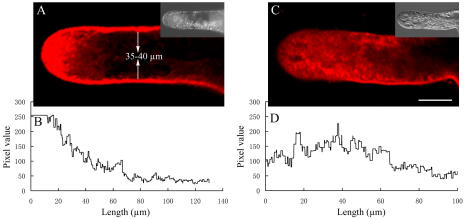
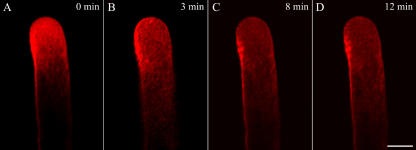
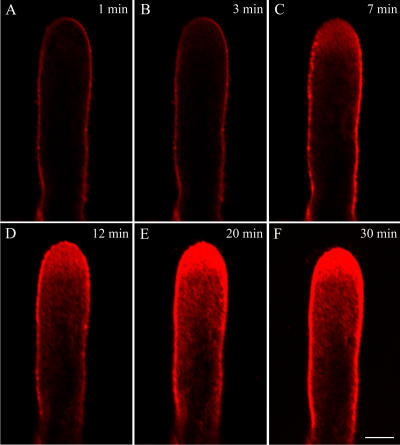
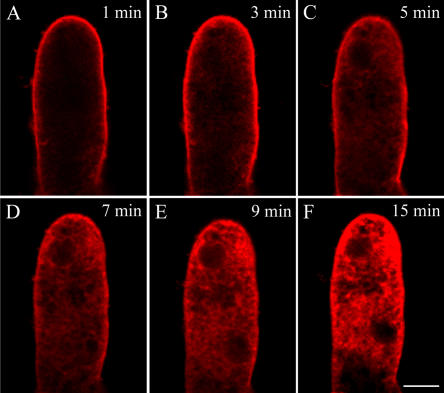
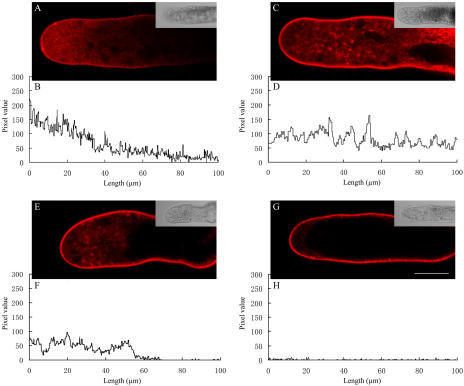
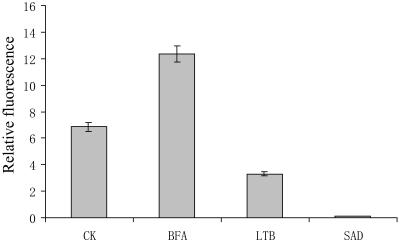
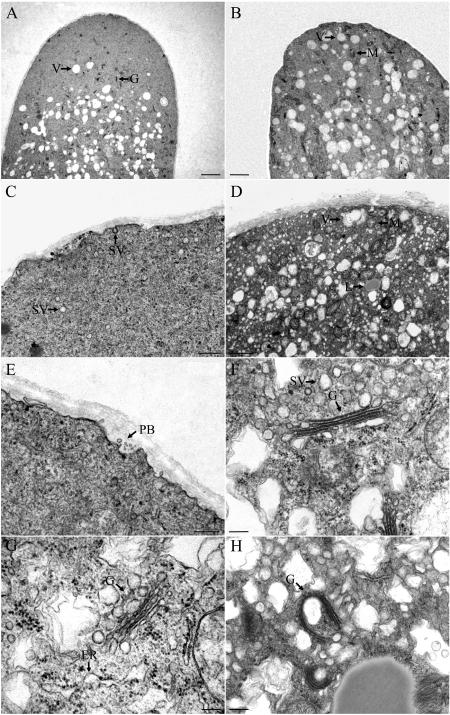
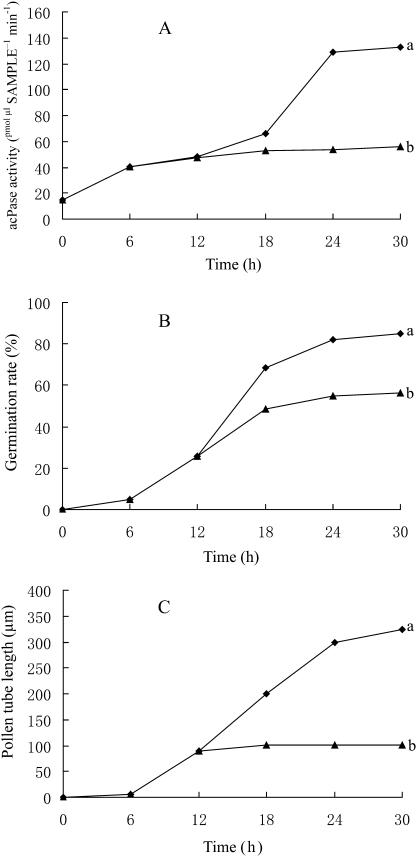
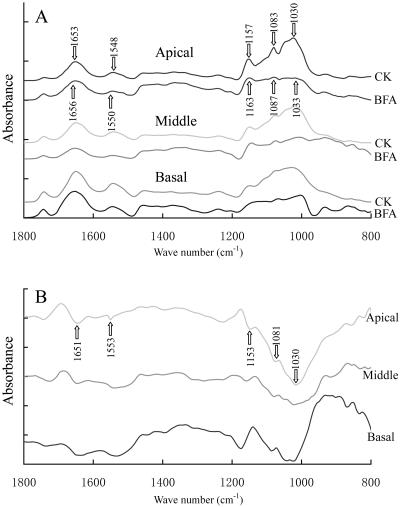
We assessed the effects of brefeldin A (BFA) on pollen tube development in Picea meyeri using fluorescent marker FM4-64 as a membrane-inserted endocytic/recycling marker, together with ultrastructural studies and Fourier transform infrared analysis of cell walls. BFA inhibited pollen germination and pollen tube growth, causing morphological changes in a dose-dependent manner, and pollen tube tip growth recovered after transferring into BFA-free medium. FM4-64 labeling showed typical bright apical staining in normally growing P. meyeri pollen tubes; this apical staining pattern differed from the V-formation pattern found in angiosperm pollen tubes. Confocal microscopy revealed that exocytosis was greatly inhibited in the presence of BFA. In contrast, the overall uptake of FM4-64 dye was about 2-fold that in the control after BFA (5 microg mL(-1)) treatment, revealing that BFA stimulated endocytosis in a manner opposite to the induced changes in exocytosis. Transmission electron microscopic observation showed that the number of secretory vesicles at the apical zone dramatically decreased, together with the disappearance of paramural bodies, while the number of vacuoles and other larger organelles increased. An acid phosphatase assay confirmed that the addition of BFA significantly inhibited secretory pathways. Importantly, Fourier transform infrared microspectroscopy documented significant changes in the cell wall composition of pollen tubes growing in the presence of BFA. These results suggest that enhanced endocytosis, together with inhibited secretion, is responsible for the retarded growth of pollen tubes induced by BFA.
Figures
References
-
- Baluška F, Liners F, Hlavacka A, Schlicht M, Van Cutsem P, McCurdy D, Menzel D (2005) Cell wall pectins and xyloglucans are internalized into dividing root cells and accumulate within cell plates during cytokinesis. Protoplasma 225: 141–155 - PubMed
-
- Bolte S, Talbot C, Boutte Y, Catrice O, Read ND, Satiat-Jeunemaitre B (2004) FM-dyes as experimental probes for dissecting vesicle trafficking in living plant cells. J Microsc 214: 159–173 - PubMed
-
- Camacho L, Malhó R (2003) Endo/exocytosis in the pollen tube apex is differentially regulated by Ca2+ and GTPases. J Exp Bot 54: 83–92 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
